Arabica coffee, native to Ethiopia, is esteemed for its exceptional quality and dominates the global specialty coffee market. As the primary cultivated coffee species, it accounts for approximately 60–65% of global coffee production. The genetic diversity of Arabica coffee, shaped through natural evolution and human domestication, is a cornerstone of its adaptability and resilience against biotic and abiotic stresses. Domestication syndrome traits such as reduced seed dispersal, compact growth, and increased uniformity have facilitated its cultivation, yet these traits have inadvertently narrowed its genetic base, making the crop more vulnerable to environmental and pathogenic threats. The genetic makeup of Arabica coffee is unique, with an allotetraploid genome that combines contributions from two diploid species, Coffea canephora and Coffea eugenioides. Despite its evolutionary significance, Arabica coffee exhibits relatively low genetic variation compared to other Coffea species. This limited diversity heightens its susceptibility to genetic erosion caused by deforestation, climate change, and unsustainable monoculture practices. Conservation efforts are crucial to preserving Arabica’s genetic resources, employing both ex-situ and in-situ strategies. Ex-situ methods include seed banks, cryopreservation, and field gene banks, while in-situ conservation protects wild populations in their natural habitats. Modern biotechnological tools such as molecular markers, genetic mapping, and somatic embryogenesis enhance the precision and efficiency of germplasm conservation and utilization. Breeding programs aim to address the challenges posed by climate change, pests, and diseases by developing varieties with enhanced drought tolerance, disease resistance, and higher yields. Hybrid vigor (heterosis) has shown promise in boosting adaptability and productivity. While vegetative propagation ensures uniformity and retention of elite traits, it limits genetic recombination, which is vital for long-term adaptability. In contrast, seed-based propagation facilitates genetic improvement but may compromise trait consistency. Notable achievements in breeding include improved cultivars like Geisha, SL28, and F1 hybrids, which balance productivity with stress resilience. Preserving Arabica coffee’s genetic base and advancing breeding efforts remain essential to securing the crop’s future and maintaining its contribution to global agriculture and livelihoods.
| Published in | International Journal of Food Science and Biotechnology (Volume 9, Issue 4) |
| DOI | 10.11648/j.ijfsb.20240904.14 |
| Page(s) | 80-106 |
| Creative Commons |
This is an Open Access article, distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution and reproduction in any medium or format, provided the original work is properly cited. |
| Copyright |
Copyright © The Author(s), 2024. Published by Science Publishing Group |
Arabica Coffee, Genetic Diversity, Germplasm Conservation, Breeding Strategies, Heterosis, Biotechnology in Coffee, Improved Coffee Varieties
No | Bourbon | Typical | Bourbon and Typical | ||
|---|---|---|---|---|---|
1 | Bourbon | 13 | Harrar Rwanda | 22 | Catuai |
2 | Bourbon Mayguez 139 | 14 | Maragogipe | 23 | Mondo Novo |
3 | Bourbon Mayguez 71 | 15 | Mibirizi | 24 | Pacamara |
4 | Caturra | 16 | Nyasaland | ||
5 | Jackson 2/1257 | 17 | Pache | ||
6 | K 7 | 18 | Pop3303/21 | ||
7 | kP423 | 19 | SL14 | ||
8 | Pacas | 20 | SL34 | ||
9 | SL28 | 21 | Typical | ||
10 | Tekisic | ||||
11 | Venecia | ||||
12 | Villa Sarchi |
25 | Geisha (Panama) |
|---|---|
26 | Java |
No | Catimor Group | Sarchimor Group | Others | ||
|---|---|---|---|---|---|
27 | Anacafe | 36 | Cuscatleco | 43 | Batian |
28 | Catimor 129 | 37 | IAPAR 59 | 44 | RAB C15 |
29 | Catisic | 38 | Limani | ||
30 | Costa Rica 95 | 39 | Marsellesa | ||
31 | IHCAFE 90 | 40 | Obata Rojo | ||
32 | Lempira | 41 | Parainema | ||
33 | Oro Azteca | 42 | T 5296 | ||
34 | T 5175 | ||||
35 | T 8667 |
No | Introgressed | Not Introgressed | |
|---|---|---|---|
45 | Centroamericano | 52 | Casiopea |
46 | Evaluna | 53 | H3 |
47 | Milenio | ||
48 | Mundo Maya | ||
49 | Nayarita | ||
50 | Ruiru 11 | ||
51 | Starmaya |
AFLP | Amplified Fragment Length Polymorphism |
CBD | Coffee Berry Disease |
CLR | Coffee Leaf Rust |
CWD | Coffee Wilt Disease |
GCA | General Combining Ability |
GMO | Genetically Modified Organisms |
MAS | Marker Assisted Selection |
PCR | Polymerization Chain Reaction |
QTL | Quantitative Trait Loci |
RAPD | Randomly Amplified Polymorphic DNA |
RFLP | Restriction Fragment Length Polymorphism |
SCA | Specific Combining Ability |
SSR | Simple Sequence Repeats |
| [1] | Adams, R. P. & Adams, J. E. (1992). Conservation of Plant Genes: DNA Banking and In vitro Biotechnology. Academic Press, San Diego, USA. |
| [2] | Adugna G, Hindorf H, Steiner U, Nirenberg H, Dehne I, et al. (2005). Genetic diversity in the coffee wilt pathogen (Gibberella xylarioides) populations: differentiation by host specialization and RAPD analysis. J Plant Dis Protect 112: 134–145. |
| [3] | Aerts RJ, Baumann TW (1994). Distribution and utilization of chlorogenic acid in Coffea seedlings. J. Exp. Bot. 45: 497-503. |
| [4] | Agwanda, C. O., Lashermes, P., Trouslot, P., Combes, M. C. and Charrier, A. (1997). Identification of RAPD markers for resistance to coffee berry disease, Colletotrichum kahawae, in arabica coffee. Euphytica 97: 241-248. |
| [5] | Allard, R. W. (1960). Principles of Plant Breeding. John Wiley and Sons. Inc. New York. |
| [6] | Ameha, M. (1983). Heterosis in crosses of indigenous coffee selected for yield and resistance to coffee berry disease. I. First bearing stage. Acta Hort. 140: 155±161. |
| [7] | Anthony, F., Combes, M. C., Astorga, C., Bertrand, B., Graziosi, G., and Lashermes, P. (2002). The origin of cultivated Coffea arabica L. varieties revealed by AFLP and SSR markers. Theor. Appl. Genet. 104: 894- 900. |
| [8] | Anthony, F.; Bertrand, B.; Quiros, O.; Lashermes, P.; Berthaud, J.; Charrier, A. (2001). Genetic diversity of wild coffee (Coffea arabica L.) using molecular markers. Euphytica 118: 53±65. |
| [9] | Bayeta, B. (2001). Arabica coffee breeding for yield and resistance to coffee berry disease (collectotrichum cahawe Sp. Nov.) A thesis submitted to the University of London in partial fulfillment of the requirements of the degree of Doctor philosophy, Wye College, university of London, U. K. |
| [10] | Bayetta, B. (1986). Exploration and coffee germplasm collection in the Gambella plain. IARNews letter, 1: 3-5. |
| [11] | Bayetta, B. (1991). Nursery evaluation of heterosis and combining ability in reference to origin and morphology of parents in coffee (Coffea arbica L.). M. Sc. Thesis. Alemaya University of Agriculture, Alemaya. 128pp. |
| [12] | Berthouly, M. and Etienne, H. (2000a). Somatic embryogenesis of coffee. p. 71-90. In: T. Sera, C. R. Soccol, A. Pandey and S. Roussos (eds.), Coffee Biotechnology and Quality. Kluwer Academic Publishers, Netherlands. |
| [13] | Berthouly, M.; Alvarand, D.; Carrasco, C. and Teisson, C. (1994). In vitro micropropagation of coffee sp. by temporary immersion. In: Abstracts: 8 International Congress of Plant Tissue and Cell Culture. Florence, Italy. p. 162. |
| [14] | Berthouly, M.; Etienne, H. (2000b). Somatic embryogenesis of Coffee. In: Jain, S. M., Gupta, P. K., Newton, R. J., eds. Somatic embryogenesis in woody plants, vol 5. London: Kluwer Academic Publishers;: 259±288. |
| [15] | Berthouly, M.; Michaux-Ferrie Á re, N. (1996). High frequency somatic embryogenesis in Coffea canephora: induction conditions and histological evolution. Plant Cell Tiss. Organ Cult. 44: 169±176. |
| [16] | Bertrand, B.; Etienne, H.; Cilas, C et al (2005). Coffea arabica hybrid performance for yield, fertility and bean weight. Euphytica 141: 255–262 2. |
| [17] | Bertrand, B.; Alpizar, E.; Llara, L.; et al (2011). Performance of Arabica F1 hybrids in agroforestry and full-sun cropping systems in comparison with pure lines varieties. Euphytica 181: 147–158. |
| [18] | Berthaud, J and Charrier A. (1998). Genetic Resources of Coffea. pp. 1-42. In Clarke RJ and Marcrae R (eds). Coffee Vol. 4 Agronomy, Elsevier Applied Science, London and New York. |
| [19] | Brooke, C. (1960). Khat (Catha edulis): its production and trade in the Middle East. Geogr J 126(1): 52–59. |
| [20] | CABI, (2003). Surveys to assess the extent and impact of coffee wilt disease in East and Central Africa, Final Technical Report, CORNET/ASARECA 36pp + 5 Appendices + 3 Annexes. |
| [21] | Carneiro, M. F. (1997). Coffee biotechnology and its application in genetic transformation. Euphytica 96: 167-172. |
| [22] | Carvalho, A. (1958). Recent advances in our knowledge of coffee trees: 2. Genetics. P. 30 - 36. In: B. Sachs (ed) Advances in crop production technology. Coffee and tea industries. |
| [23] | Carvalho, A. and L. C. Monaco. (1962). Natural cross pollination in Coffea arabica L. Proc. 16th Int. Hort. Congr. 4: 447 - 449. |
| [24] | Carvalho, A. (1988). Principles and practice of coffee plant breeding for productivity and quality factors: Coffea arabica. In: Fewerda, F. P., Wit, F., eds Coffee: agronomy, vol. 4. London: Elsevier Applied Science: 129±165. |
| [25] | Carvalho, A., F. P.; Ferwerda, J. A.; Frahm-lelived, D. M.; Medina and L. C. Monaco. (1969). Coffee (C. arabica L and C. canephora Pierre ex Froehner). P. 186-244. In: F. P. Ferwerda and F. Wilt (eds) Outlines of perennial crop breeding in the tropics. Landbouwhogeschool (Agricultural University) Wageningen, The Netherlands. |
| [26] | Charrier, A and Eskes, AB. (2004). Botany and Genetics of coffee. pp. 25-55. In Wintgens JN (ed). Coffee: growing, processing, sustainable production. Wiley. VCH, Weiheim. |
| [27] | Charrier, A. (1977). La structure génétique des caféiers spontanés de la région Malgache (Mascarocoffea). Leurs relations avec les caféiers d’origine africaine (Eucoffea). Mémoires ORSTOM 87, IRD, Paris. |
| [28] | Charrier, A. (1978). Etude de la structure et de la variabilite´ ge´ne´tique des cafe´iers. Bulletin IFCC 14: 100. |
| [29] | Charrier, A. and Berthaud, J. (1985). Botanical classification of coffee. In: Clifford, M. N. and Wilson, K. C.(eds), Coffee Botany, Biochemistry and Production of Beans and Beverage, Croom Helm, London, Pp. 13-47. |
| [30] | Custer, J. B. M.; Van Ed., G. and Buijs, L. C. (1980). Clonal propagation of Coffea Arabica L. by nodal culture. IX Intl in vitro Scientific Colloquium on Coffee, London, ASIC, Paris, p. 589-596. |
| [31] | Davis, A. P.; Gole, T. W.; Baena, S., & Moat, J. (2012). The impact of climate change on indigenous Arabica coffee (Coffea arabica): Predicting future trends and identifying priorities. PLoS ONE, 7(11), e47981. |
| [32] | De Brito, G. G.; Caixeta, E. T.; Gallina, A. P. et al. (2010). Inheritance of coffee leaf rust resistance and identification of AFLP markers linked to the resistance gene. Euphytica 173: 255-264. |
| [33] | De Kochko, A.; Akaffou, S.; Andrade, A. C. et al. (2010). Advances in Coffee Genomics. In: J.-C. Kader and M. Delseny (eds.), Advances in Botanical Research (Volume 53). Academic Press. |
| [34] | Dessalegn, Y.; Herselman, L. and Labuschagne, M., (2009). Comparison of SSR and AFLP analysis for genetic diversity assessment of Ethiopian arabica coffee genotypes. South African Journal of Plant and Soil, 26(2), pp. 119-125. |
| [35] | Dufour, M.; Leroy, T.; Carasco-Lacombe, C.; Philippe, R. and Fenouillet, C. (2000). Coffee (Coffea sp.) genetic transformation for insect resistance. p. 209-217. In: T. Sera, C. R. Soccol, A. Pandey and S. Roussos (eds.), Coffee Biotechnology and Quality. Kluwer Academic Publishers, Netherlands. |
| [36] | Dulloo, M. E.; Charrier, A.; Dussert, S.; Anthony, F.; Tesfaye, S.; Rakotomalala, J. J. & Agwanda, C. (2001). Conservation of coffee genetic resources: constraints and opportunities. Ref. B208, in: Proceedings (CD-ROM) of the 19th International Scientific Colloquium on Coffee. Trieste, Italy, 14–18 May 2001. ASIC, Montpellier, France. |
| [37] | Dulloo, M. E.; Guarino, L.; Engelmann, F. et al. (1998). Complementary conservation strategies for the genus Coffea: A case study of Mascarene Coffea species. Genet. Resour. Crop Evol. 45: 565-579. |
| [38] | Dussert, S.; Chabrillange, N.; Anthony, F.; Engelmann, F.; Recalt, C. & Hamon, S. (1997a). Variability in storage response within a coffee (Coffea spp.) core collection under slow-growth conditions. Plant Cell Reports 16: 344–348. |
| [39] | Dussert, S.; Chabrillange, N.; Engelmann, F.; Anthony, F. & Hamon, S. (1997b). Cryopreservation of coffee (Coffea arabica L.) seeds: importance of the precooling temperature. CryoLetters 18: 269–276. |
| [40] | Dussert, S.; Chabrillange, N.; Vasquez, N.; Engelmann, F.; Anthony, F.; Guyot, A. & Hamon, S. (2000). Beneficial effect of post-thawing osmoconditioning on the recovery of cryopreserved coffee (Coffea arabica L.) seeds. CryoLetters 21: 47–52. |
| [41] | Dussert, S.; Engelmann, F.; Chabrillange, N.; Anthony, F.; Noirot, M. & Hamon, S. (1997c). In vitro conservation of coffee (Coffea spp.) germplasm. pp. 287–305, in: M. K. Razdan and E. C. Cocking (editors). Conservation of Plant Genetic Resources In Vitro Vol. 1. M/S Science, USA. |
| [42] | Dussert, S.; Vasquez, N.; Salazar, K.; Anthony, F. and Engelmann, F. (2007). Cryopreservation of coffee genetic resources. p. 49-58. In: F. Engelmann, M. E. Dulloo, C. Astorga, S. Dussert and F. Anthony (eds.), Complementary strategies for ex situ conservation of coffee (Coffea arabica L.) genetic resources. A case study in CATIE, Costa Rica. Tropical reviews in Agricultural Biodiversity. Bioversity International, Rome, Italy. |
| [43] | Duvic, D. N. (1997). Commercial strategies for exploiting heterosis. P. 206 - 207. In: CIMMYT books of abstracts, the genetics and exploitation of heterosis in crops: an international symposium. Mexico, D. F., Mexico. |
| [44] | Eskes, A. B. & Toma – Braghinin M. (1981). Assessment methods for resistance to coffee leaf rust, Hemileia vastatix Bark & Br. Plant Protection Bulletin. FAO 29: 56 -56. |
| [45] | Etienne, H.; Anthony, F.; Dussert, S.; Fernandez, D.; Lashermes, P. & Bertrand, B. (2002). Biotechnological applications for the improvement of coffee (Coffea arabica L.). In Vitro Cellular and Development Biology - Plant 38: 129–138. |
| [46] | Etienne, H.; Berthouly, M. (2002). Temporary immersion systems in plant micropropagation (review). Plant Cell Tiss. Organ Cult. (in press). |
| [47] | Etienne, H.; Bertrand, B.; Anthony, F.; Co à te, F.; Berthouly, M. (1997). Somatic embryogenesis: a tool for coffee breeding. Proceedings of 17th Colloquium of International Coffee Science Association, Nairobi, Kenya. Vevey, Switzerland: ASIC;: 457±465. |
| [48] | Ferwerda, F. P. (1986). Coffee: coffee species (Rubiaceae). P. 231 - 249. In: L. C. A. Leakey (ed.) Crop improvement in East Africa. Billing and Sons Ltd, Guildford and London. |
| [49] | Friis, I.; Demissew, S. & Breugel, P. V. (2010). Atlas of the potential vegetation of Ethiopia. Biol. Skrif. 58, 1–307. |
| [50] | Gichuru, E. K.; Agwanda, C. O.; Combes, M. C. et al. (2008). Identification of molecular markers linked to a gene conferring resistance to coffee berry disease (Colletotrichum kahawae) in Coffea arabica. Plant Pathol. 57: 1117-1124. |
| [51] | Girma A,; Hulluka M, & Hindorf H. (2001). Incidence of tracheomycosis, Gibberella xylarioides (Fusarium xylarioides) on Arabica coffee in Ethiopia. J Plant Dis Prot 108(2): 136– 142. |
| [52] | Gole, T. W. (2003). Conservation and Use of Coffea Genetic Resources in Ethiopia: Challenges and Opportunities in the Context Current Global Situations. |
| [53] | Gole, T. W.; Denich, M.; Teketay, D. and Borsch, T. (2001). Diversity of traditional coffee production systems in Ethiopia and their contributions to the conservation of coffee genetic diversity. Conference on International Agricultural Research for Developpment. Deutscher Tropentag, Bonn, 9-11 October. |
| [54] | Gole, T. W.; Denich, M.; Teketay, D. Vlek, P. L. G. (2002). Human impacts on Coffea arabica gene pools in Ethiopia and the need for its in situ conservation. In: R. Rao, A. Brown, M. Jackson (eds.) Managing Plant Genetic Diversity. CABI and IPGRI, 237-247. |
| [55] | Haarer, A. E. (1962). Modern Coffee Production, 2nd ed. London. |
| [56] | Hatanaka, T.; Choi, YE.; Kusano T.; Sano, H. (1999). Transgenic plants of Coffea canephora from embryogenic callus via Agrobacterium tumefaciens-mediated transformation. Plant Cell Reports 19, 106-110. |
| [57] | Hayes, J. D. and C. A. Foster. (1976). Heterosis in self-pollinated crops, with particular reference to barley. P239 - 256. In: A. Janossy and F. G. H. Lupton (eds) Heterosis in plant breeding, proceedings of the 7th congress of EUCARPIA. Elsevier Scientific Publishing Company, Amsterdam. |
| [58] | Hayman, B. I. (1960). Heterosis and quantitative inheritance. Heredity, 15: 324-327. |
| [59] | Hein, L & Gatzweiler, F. (2006). The economic value of coffee (Coffea arabica) genetic resources. Ecol Econ 60: 176–185. |
| [60] | Herrera, J. C.; Combes, M. C.; Anthony, F.; Charrier, A. and Lashermes, P. (2002). Introgression into the allotetraploid coffee (Coffea arabica L.): segregation and recombination of the C. canephora genome in the tetraploid interspecific hybrid (C. arabica x C. canephora). Theor. Appl. Genet. 104: 661-668. |
| [61] | Herrera, P. J. C.; Alvarado, A. G.; Cortina, G. H. A.; Combes, M. C.; Romero, G. G. and Lashermes, P. (2009). Genetic analysis of partial resistance to coffee leaf rust (Hemileia vastatrix Berk & Br.) introgressed into the cultivated Coffea arabica L. from the diploid C. canephora species. Euphytica 167: 57-67. |
| [62] | IPGRI (1996). Description for Coffee (Coffea sp. and Psilanthus sp.). International Plant Genetic Resource Institute, Rome. |
| [63] |
IUCN, UNEP (2010). The World Database on Protected Areas (WDPA). UNEPWCMC. Cambridge, UK. Available:
www.protectedplanet.net Accessed 2012 Jan 3. |
| [64] | Johnson, R. (1992). Past, present and future opportunities in breeding for disease resistance, with examples from wheat. Euphytica 63: 3 - 22. |
| [65] | Jury, M. R. (2014). Statistical evaluation of CMIP5 climate change model simulations for the Ethiopian highlands. Int. J. Climatol. 35, 37–44. |
| [66] | Krug, C. A. and A. Carvalho. (1952). Coffee breeding. Rep. 13th Int. Hort. Congr., Brussels. P. 2 - 33. |
| [67] | Krug,. A. & Carvalho, A. (1951). The genetics of Cojfea. Advances in Genetics, New York, 4: 127-158. |
| [68] | Kumar, V.; Naidu, M. M. and Ravishankar, G. A. (2006). Developments in coffee biotechnology – in vitro plant propagation and crop improvement. Plant Cell Tiss. Organ Cult. 87: 49-65. |
| [69] | Labouisse, J-P.; Bellachew, B.; Kotecha, S. & Bertrand B. (2008). Current status of coffee (Coffea arabica L.) genetic resources in Ethiopia: implications for conservation. Genet Resour Crop Ev 55: 1079–1093. |
| [70] | Lashennes, P.; Trouslot, P.; Anthony, F.; Combes, M. C & Charrier, A. (l996a). Genetic diversity for RAPD markers between cultivated and wild accessions of Coffea arabica. Euphytica 87: 59 - 64. |
| [71] | Lashermes, P.; Combes, M. C.; Topart, P.; Graziosi, G.; Bertrand, B. and Anthony, F. (2000b). Molecular breeding in coffee (Coffea arabica L.). p. 101-112. In: T. Sera, C. R. Soccol, A. Pandey and S. Roussos (eds.), Coffee Biotechnology and Quality. Kluwer Academic Publishers, Netherlands. |
| [72] | Lashermes, P.; Cros, J.; Combes, M. e.; Trouslot, P.; Anthony, F., Hamon, S. & Charrier, A. (1996a), Inheritance and restriction fragment lenght polymorphism of chloroplast DNA in the genus Cojfea L. Theor. App\. Genet._93: 626 -632. |
| [73] | Lashermes, P. J., Andrzejewiski., Bertrand, B., Combes, M. C., Dussert, S., Graziosi, G., Trouslot, P. and Anthony, F. (2000a). Molecular analysis of introgressive breeding in coffee (Coffea abica L.). Theor. Appl. Genet. 100: 139-146. |
| [74] | Leroy, T.; Henry AM.; Royer M et al (2000). Genetically modified coffee plants expressing the Bacillus thuringiensis cry 1Ac gene for resistance to leaf miner. Plant Cell Rep 19: 382–389. |
| [75] | Li, J. and Yuan, L. (2000). Hybrid rice: Genetics, breeding and seed production. Plant Breed. Rev. 17: 15 — 158. |
| [76] | Mackey, J. (1976). Genetic and evolutionary principles of heterosis. P. 17 - 32. In; A. Janossy and F. G. H. Lupton (eds) Heterosis in plant breeding, proceedings of the 7th congress of EUCARPIA. Elsevier Scientific publishing company, Amsterdam. |
| [77] | Mayo, O. (1980). The Theory of Plant Breeding. Oxford University Press, New York. 293 pp. |
| [78] | Mayo, O. (1987). The theory of plant breeding. Clarendon Press, Oxford. 334pp. |
| [79] | McSweeney, C., New, M. & Lizcano, G. (2010). UNDP Climate Change Country Profiles: Ethiopia (UNDP, 2010); |
| [80] | Medina Filho, H. P.; Carvalho, A.; Sondahl, M. R.; Fazuoli, L. C. & Costa, W. M. (1984). Coffee breeding related evolutionary aspects. In: Plant Breeding Reviews (J. Janick, ed.). Avi Publish. Co. Connecticut, USA V. 2 p. 157-193. |
| [81] | Mendes AJT. (1941). Cytological observations in Coffea VI. Embryo and endosperm development in Coffea arabica L. Am. J. Bot. 28: 784-789. |
| [82] | Mesfin, A. & Bayetta, B. (1987). Genotype-Environmental interactions in coffee (Coffea arabica L.). 12th. International scientific colloQuium on coffee (ASIC). Paris pp. 476482. |
| [83] | Mesfin, A. (1982). Heterosis in crosses of indigenous coffee (C. arabica L.) selected for yield and resistance to coffee berry disease: I. At first bearing stage. Eth. J. Agr. Sci., IV: 33 -43. |
| [84] | Mesfin, A. and Bayetta, B. (1983). Heterosis in crosses of indigenous coffee selected for yield and resistance to coffee berry disease: II. First three years. Eth. J. Agr. Sci., V: 13 -21. |
| [85] | Meyer, FG.; Fernie LM.; Narasimhaswamy RL.; Monaco LC.; Greathead DJ. (1968). FAO coffee mission to Ethiopia 1964–1965. FAO, Rome. |
| [86] | Meyer, F. G. (1965). Notes on wild Coffee arabica from South western Ethiopia, with some historical considerations. Econ. Bot. 19: 136 – 151. |
| [87] | Moat, J.; Gole, TW. & Davis AP. (2019). Least concern to endangered: Applying climate change projections profoundly influences the extinction risk assessment for wild Arabica coffee. Glob Change Biol. 2019; 00: 1–14. |
| [88] | Moat, J.; Williams, J.; Baena, S.; Wilkinson, T.; Gole, T. W.; Challa, Z. K. & Davis, A. P. (2017). Resilience potential of the Ethiopian coffee sector under climate change. Nature Plants, 3, 17081. |
| [89] | Monaco, L. C. (1968). Considerations on the genetic variability of the Coffea arabica populations in Ethiopia. In: FAO (Eds), FAO Coffee Mission to Ethiopia 1964-65, FAO, Rome, Italy. Pp 49 – 69. |
| [90] | Motta, L. B.; Soares, T. C. B.; Ferrão, M. A. G.; Caixeta, E. T.; Lorenzoni, R. M. and Souza Neto, J. D. D. (2014). Molecular characterization of Arabica and Conilon coffee plants genotypes by SSR and ISSR markers. Brazilian archives of biology and technology, 57(5), pp. 728-735. |
| [91] | Musoli, P.; Cubry, P.; Aluka, P. et al. (2009). Genetic differentiation of wild and cultivated populations: diversity of Coffea canephora Pierre in Uganda. Genome 52: 634-646. |
| [92] | N’Diaye, A.; Noirot, M.; Hamon, S. and Poncet, V. (2007). Genetic basis of species differentiation between Coffea liberica Hiern and C. canephora Pierre: Analysis of an interspecific cross. Genet. Resour. Crop Evol. 54: 1011-1021. |
| [93] | Noir, S.; Anthony, F.; Bertrand, B.; Combes, M. C. and Lashermes, P. (2003). Identification of a major gene (Mex-1) from Coffea canephora conferring resistance to Meloidogyne exigua in Coffea arabica. Plant Pathol. 52: 97-103. |
| [94] | Orozco-Castillo, C.; Chalmers, K. J.; Waugh, R. and Powell, W. (1994). Detection of genetic diversity and selective gene introgression in coffee using RAPD markers. TAG Theoretical and Applied Genetics, 87(8), pp. 934-940. |
| [95] | Osorio, N. (2002). The global coffee crisis: a threat to sustainable development. ICO report. Submitted to the World Summit on Sustainable Development, Johannesburg. Available at |
| [96] | Ovalle-Rivera, O.; Läderach, P.; Bunn, C.; Obersteiner, M. & Schroth, G. (2015). Projected shifts in coffea arabica suitability among major global producing regions due to climate change. PLoS ONE 10, e0124155. |
| [97] | Paulos, D, Demil, T. (2000). The need for forest coffee germplasm conservation in Ethiopia and its significance in the control of coffee diseases. In: Proceedings of the workshop on control of Coffee Berry Disease (CBD) in Ethiopia held in Addis Ababa. Ethiopian Agricultural Research Organization, Addis Ababa, pp 125–134. |
| [98] | Pearl, H. M. C.; Nagai, P. H.; Moore, D. L.; Steiger, R. V.; Osgood and Ming, R. (2004). Construction of a genetic map for arabica coffee. Theor. and Appl. Genet. 108: 829-835. |
| [99] | Pendergrast, M. (2009). Coffee second only to oil? Tea & Coffee Trade Journal. April: 38- 41. |
| [100] | Philippe, L.; Benoȋt, B. and Harvé, E. (2009). Breeding coffee (Coffea arabica L) for sustainable production. In: S. M. Jain and P. M. Priyadarshan (eds.), Breeding Plantation Tree Crops: Tropical Species. Springer Science+Business Media. LLC. |
| [101] | Powell, W.; Orozco-Castillo, C.; Chalmers, K. J.; Provan, J. and Waugh, R. (1995). Polymerase chain reaction-based assays for the characterisation of plant genetic resources. Electrophoresis, 16(1), pp. 1726-1730. |
| [102] | Prakash, N. S.; Combes, M. C.; Somanna, N. and Lashermes, P. (2002). AFLP analysis of introgression in coffee cultivars (Coffea arabica L.) derived from a natural interspecific hybrid. Euphytica 124: 265-271. |
| [103] | Raina, S. N.; Mukai, Y. and Yamamoto, M. (1998). In situ hybridisation identifies the diploid progenitor species of Coffea arabica (Rubiaceae) (abstract). Theoret. Appl. Genet. 97: 8. |
| [104] | Rhodes, G.; Ju, W.; Yang Ju and Samaras Y. (1992). Plant metabolism and heterosis. Plant Breed. Rev. 10: 54-9 1. |
| [105] | Santana-Buzzy, N.; Rojas-Herrara, R.; Galaz-Avalos, R. M.; Ku-Cauich, J. R.; MijangosCortes, J.; Gutierrez-Pancheco, L. C.; Canto, A.; Quiroz-Figueroa, F. and LoyolaVargas, V. M. (2007). Advances in coffee tissue culture and its practical applications. In Vitro Cell. Dev. Biol. Plant. 43: 507-520. |
| [106] | Senbeta, F. and Denich, M. (2006). Effects of wild coffee management on species Diversity in the afromontane rain forest of Ethiopia. Forest Ecology and Management. 232: 68–74. |
| [107] | Senbeta, F. (2006). Biodiversity and Ecology of Afromontane Rainforests with Wild Coffea arabica L. Populations in Ethiopia: PhD thesis, Ecology and Development Sreries, No 38, Cuvillier Verlag Göttingen, University of Bonn, Germany. No. 38. |
| [108] | Sharp, W. R.; Caldas, L. S.; Crocomo, O. J.; Monaco, L. C. and Carvalho, A. (1973). Production of Coffea arabica callus of three ploidy levels and subsequent morphogenesis. Phyton. 31: 67-74. |
| [109] | Söndahl, M. R.; Nakamura, T.; Medina-Filho, H. P.; Carvalho, A.; Fazuoli, L. C. and Costa, W. M. (1984). Coffee. p. 564-590. In: P. V. Ammirato et al. (eds.), Handbook of Plant Cell Culture, Crop Species, Vol. 3, McMillan, New York. |
| [110] | Söndahl, M. R.; Söndahl, C. N. and Goncalves, W. (2000). Field testing of Arabica bioreactor-derived plants. p. 143-150. In: T. Sera, C. R. Soccol, A. Pandey and S. Roussos (eds.), Coffee Biotechnology and Quality. Kluwer Academic Publishers, Netherlands. |
| [111] | Spiral, J.; Thierry, C.; Paillard, M. and Petiard, V. (1993). Obtention de plantules de Coffea canephora Pierre (Robusta) transformées par Agrobacterium rhizogenes. C. R. Acad. Sci. Paris 316. Série III: 1-6. |
| [112] | Spiral, J.; Leroy, T.; Paillard, M. & Pe´tiard, V. (1999). Transgenic coffee (Coffea species). In: Bajaj, Y. P. S., ed. Biotechnology in agriculture and forestry, vol. 44. Transgenic trees. Berlin, Heidelberg: SpringerVerlag; 55–76. |
| [113] | Sridevi, V.; Giridhar, P.; Simmi, P. S. and Ravishankar, G. A. (2010). Direct shoot organogenesis on hypocotyls explants with collar region from in vitro seedlings of Coffea canephora Pierre ex. Frohner cv. C x R and Agrobacterium tumefaciensmediated transformation. Plant Cell Tiss. Organ Cult. 101: 339-347. |
| [114] | Srinivasan, C. S. and Vishveshwara. S. (1978). Heterosis and stability for yield in arabica * coffee. Indian J. Genet., PI. breed. 38: 13 -21. |
| [115] | Srivastava, H. K. (1981). Intergenomic interaction, heterosis and improvement of crop yield. Adav. Agron. 34: 117 - 195. |
| [116] | Stuber, C. W. (1997). The biology and physiology of heterosis. P. 108-9. In: CIMMYT books of abstracts, the genetic and exploitation of heterosis in crops: an international symposium. Mexico, D. F., Mexico. |
| [117] | Sugiyama, M.; Matsuoka, C.; Takagi, T. (1995). Transformation of coffee with Agrobacterium rhizogenes. Proceedings of the XVI Scientific Colloquium on Coffee, ASIC, Kyoto, Japan. Paris: ASIC: 853–859. |
| [118] | Sybenga, J. (1960). Genetics and cytology of coffee: A literature review. Bibliographica Genetica XIX: 217 -316. (80). |
| [119] | Sylvain, P. G. (1958). Ethiopian Coffee: Its significance to world coffee problems. Botany 12: 111-139. |
| [120] | Taye, K. and Jurgen, (2008). Ecophysiolog of wild coffee population in mountane rain forest welgekno of Ethiopia. Proceedings of coffee knowledge and diversity. work shop of EIAR. Addis Abeba. |
| [121] | Tewolde BG. (1990). The importance of Ethiopian forests in the conservation of Arabica coffee gene-pools. In: Proceedings of the 12th plenary meeting on AETFAT, Hamburg, 4–10 September 1988, pp 65–72. |
| [122] | Va´squez, N.; Dussert, S.; Salazar, K.; Anthony, F. & Engelmann, F. (2001). Variability in response of seeds to cryopreservation within a coffee core collection. Final Report. Rome: IPGRI;: 14 pp. |
| [123] | Van Boxtel, J.; Berthouly, M.; Carasco, C.; Dufour, M.; Eskes, A. (1995). Transient expression of beta-glucuronidase following biolistic delivery of foreign DNA into coffee tissues. Plant Cell Rep. 14: 748–752; |
| [124] | Van der Graaf, N. (1981). Selection of Arabica coffee types resistant to Coffee Berry Disease in Ethiopia. Mededelingen Landbouwhogescholan, Wageningen, The Netherlands. |
| [125] | Van der Vossen, H. A. M. (1985). Coffee selection and breeding. P. 49 - 96. In: M. N. Clifford and K. C. Willson (eds) Coffee: Botany, Biochemistry and production of beans and beverages. Croom Helm Ltd, London. |
| [126] | Van der Vossen, H. A. M. and Walyaro. D. J. A. (1981). The coffee breeding program in Kenya: A review of progress made since 1971 and plan of action for the coming years. Kenya coffee, 46: 113 - 130. |
| [127] | Vavilov, N. I. (1951). The origin, variation, immunity and breeding of cultivated plants (Vol. 72, No. 6, p. 482). LWW. |
| [128] | Vega, F. E.; Ebert, A. W. and Ming, R. (2008). Coffee germplasm resources, genomics, and breeding. p. 415-447. In: J. Janick (ed.), Plant Breeding Reviews. John Wiley & Sons, Inc. |
| [129] | Walyaro. D. J. A (1983). Consideration in breeding of improved yield and Quality in Arabica coffee (Coffea arabica L.) Wagengen, the Neitherlands. |
| [130] | Walyaro, D. J. & van der Vossen, H. A. M. (1977). Pollen longevity and artificial cross-pollination in Coffea arabica L. Euphytica 26: 225–231. |
| [131] | Weising, K.; Nybom, H.; Wolff, K. and Kahl, G. (2005). DNA Fingerprinting in Plants: Principles, Methods, and Applications. Second Edition. CRC Press, Boca Raton, FL. |
| [132] | Welsh, J. R. (1990). Fundamentals of Plant Genetics and Breeding. Robert E. Krieger Publishing Company. Malbara, Florida. 290 pp. 32Breeding. Robert E. Krieger Publishing Company. Malbara, Florida. 290 pp. |
| [133] | Yeshitila, K. (2004). Prospects of forest genetic resources conservation, In: Proceeding of the public meeting on integrated forest policy Dervelopment in Ethiopia, Forum for Environment (FfE), Institute of biodiversity, Addis Ababa. |
| [134] | Zirkle, C. (1952). Early ideas on breeding and cross breeding. P. l - 14. In: J. W. Gowen (ed.) Heterosis. Iowa State College Press, Ames. |
APA Style
Nasiro, K. (2024). Arabica Coffee: Genetic Diversity, Conservation Challenges, and Breeding Approaches. International Journal of Food Science and Biotechnology, 9(4), 80-106. https://doi.org/10.11648/j.ijfsb.20240904.14
ACS Style
Nasiro, K. Arabica Coffee: Genetic Diversity, Conservation Challenges, and Breeding Approaches. Int. J. Food Sci. Biotechnol. 2024, 9(4), 80-106. doi: 10.11648/j.ijfsb.20240904.14
AMA Style
Nasiro K. Arabica Coffee: Genetic Diversity, Conservation Challenges, and Breeding Approaches. Int J Food Sci Biotechnol. 2024;9(4):80-106. doi: 10.11648/j.ijfsb.20240904.14
@article{10.11648/j.ijfsb.20240904.14,
author = {Kalifa Nasiro},
title = {Arabica Coffee: Genetic Diversity, Conservation Challenges, and Breeding Approaches
},
journal = {International Journal of Food Science and Biotechnology},
volume = {9},
number = {4},
pages = {80-106},
doi = {10.11648/j.ijfsb.20240904.14},
url = {https://doi.org/10.11648/j.ijfsb.20240904.14},
eprint = {https://article.sciencepublishinggroup.com/pdf/10.11648.j.ijfsb.20240904.14},
abstract = {Arabica coffee, native to Ethiopia, is esteemed for its exceptional quality and dominates the global specialty coffee market. As the primary cultivated coffee species, it accounts for approximately 60–65% of global coffee production. The genetic diversity of Arabica coffee, shaped through natural evolution and human domestication, is a cornerstone of its adaptability and resilience against biotic and abiotic stresses. Domestication syndrome traits such as reduced seed dispersal, compact growth, and increased uniformity have facilitated its cultivation, yet these traits have inadvertently narrowed its genetic base, making the crop more vulnerable to environmental and pathogenic threats. The genetic makeup of Arabica coffee is unique, with an allotetraploid genome that combines contributions from two diploid species, Coffea canephora and Coffea eugenioides. Despite its evolutionary significance, Arabica coffee exhibits relatively low genetic variation compared to other Coffea species. This limited diversity heightens its susceptibility to genetic erosion caused by deforestation, climate change, and unsustainable monoculture practices. Conservation efforts are crucial to preserving Arabica’s genetic resources, employing both ex-situ and in-situ strategies. Ex-situ methods include seed banks, cryopreservation, and field gene banks, while in-situ conservation protects wild populations in their natural habitats. Modern biotechnological tools such as molecular markers, genetic mapping, and somatic embryogenesis enhance the precision and efficiency of germplasm conservation and utilization. Breeding programs aim to address the challenges posed by climate change, pests, and diseases by developing varieties with enhanced drought tolerance, disease resistance, and higher yields. Hybrid vigor (heterosis) has shown promise in boosting adaptability and productivity. While vegetative propagation ensures uniformity and retention of elite traits, it limits genetic recombination, which is vital for long-term adaptability. In contrast, seed-based propagation facilitates genetic improvement but may compromise trait consistency. Notable achievements in breeding include improved cultivars like Geisha, SL28, and F1 hybrids, which balance productivity with stress resilience. Preserving Arabica coffee’s genetic base and advancing breeding efforts remain essential to securing the crop’s future and maintaining its contribution to global agriculture and livelihoods.
},
year = {2024}
}
TY - JOUR T1 - Arabica Coffee: Genetic Diversity, Conservation Challenges, and Breeding Approaches AU - Kalifa Nasiro Y1 - 2024/12/13 PY - 2024 N1 - https://doi.org/10.11648/j.ijfsb.20240904.14 DO - 10.11648/j.ijfsb.20240904.14 T2 - International Journal of Food Science and Biotechnology JF - International Journal of Food Science and Biotechnology JO - International Journal of Food Science and Biotechnology SP - 80 EP - 106 PB - Science Publishing Group SN - 2578-9643 UR - https://doi.org/10.11648/j.ijfsb.20240904.14 AB - Arabica coffee, native to Ethiopia, is esteemed for its exceptional quality and dominates the global specialty coffee market. As the primary cultivated coffee species, it accounts for approximately 60–65% of global coffee production. The genetic diversity of Arabica coffee, shaped through natural evolution and human domestication, is a cornerstone of its adaptability and resilience against biotic and abiotic stresses. Domestication syndrome traits such as reduced seed dispersal, compact growth, and increased uniformity have facilitated its cultivation, yet these traits have inadvertently narrowed its genetic base, making the crop more vulnerable to environmental and pathogenic threats. The genetic makeup of Arabica coffee is unique, with an allotetraploid genome that combines contributions from two diploid species, Coffea canephora and Coffea eugenioides. Despite its evolutionary significance, Arabica coffee exhibits relatively low genetic variation compared to other Coffea species. This limited diversity heightens its susceptibility to genetic erosion caused by deforestation, climate change, and unsustainable monoculture practices. Conservation efforts are crucial to preserving Arabica’s genetic resources, employing both ex-situ and in-situ strategies. Ex-situ methods include seed banks, cryopreservation, and field gene banks, while in-situ conservation protects wild populations in their natural habitats. Modern biotechnological tools such as molecular markers, genetic mapping, and somatic embryogenesis enhance the precision and efficiency of germplasm conservation and utilization. Breeding programs aim to address the challenges posed by climate change, pests, and diseases by developing varieties with enhanced drought tolerance, disease resistance, and higher yields. Hybrid vigor (heterosis) has shown promise in boosting adaptability and productivity. While vegetative propagation ensures uniformity and retention of elite traits, it limits genetic recombination, which is vital for long-term adaptability. In contrast, seed-based propagation facilitates genetic improvement but may compromise trait consistency. Notable achievements in breeding include improved cultivars like Geisha, SL28, and F1 hybrids, which balance productivity with stress resilience. Preserving Arabica coffee’s genetic base and advancing breeding efforts remain essential to securing the crop’s future and maintaining its contribution to global agriculture and livelihoods. VL - 9 IS - 4 ER -
Department of Coffee Agronomy, Institute Ethiopian Agricultural Research, Jimma Agricultural Research Center, Jimma, Ethiopia
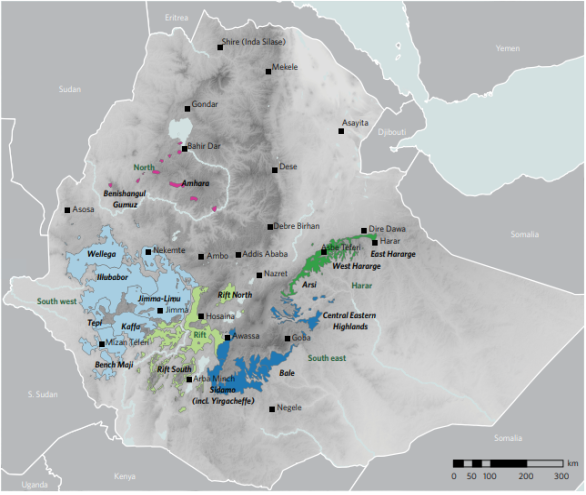
Figure 1. Map of potential wild Arabica coffee in Ethiopia and South Sudan. The colored areas represent the coverage of the humid forest types* where Light Blue represents South West coffee areas: Wellega, Illubabor, Jimma, Kaffa, Tepi and Bench Maji; LIGHT GREEN, represents Rift coffee areas i.e., Rift North and Rift South; DARK BLUE represents South East coffee areas: Sidamo, Yirgacheffe, Bale and Central Eastern Highlands; DARK GREEN represents Harar coffee areas: Arsi, West Hararge and East Hararge and PINK represents the North coffee areas: Amhara and Benishangul Gumuz. Wild Arabica coffee could occur (where there is =1% of forest cover in each km2). Map generated from species distribution models (SDMs) and remote sensing [31] [one SDM]; [88] [SDMs and remote sensing]). *Humid forest represented by Moist Evergreen Afromontane Forest (MAF) and Transitional Rain Forest (TRF) types [49]. Agroforestry systems in Sidama (south of Hawassa) are no longer wild habitats but may contain wild type plants originating from this area. Other forest areas may be highly modified compared to primary forest areas.
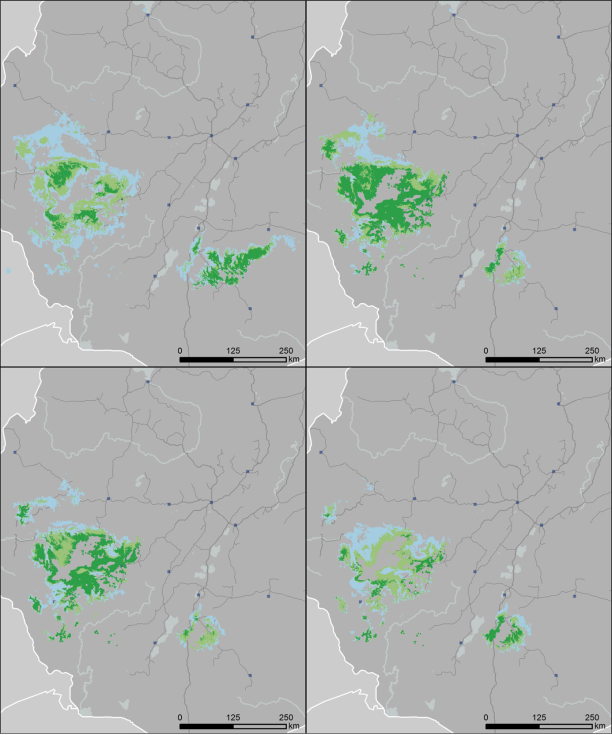
Figure 2. Maps and metrics for one example future projection; emission scenario A1b, GCM gfdl_cm2_1 and migration scenario D (see Table 1) showing SDMs and figures for AOO, EOO, and population numbers, for 1960–1990, 2010–2039, 2040–2069, and 2070–2099. The record from Bahir Dar (in the far north, for the time periods 1960–1990 and 2010–2039) is included here, although it is uncertain whether this represents an indigenous population [31].

Figure 3. Picture showing the flower and fruit of Arabica coffee.
Information

