Microsatellite instability (MSI) is a hallmark of mismatch repair (MMR) deficiency and characterizes a distinct subset of colorectal cancers (CRC). In parallel, telomere length dynamics have emerged as important contributors to genomic stability and tumorigenesis. However, the relationship between MSI status, MMR protein expression, and telomere maintenance remains poorly defined. This study aimed to investigate the association between MSI status and telomere length in CRC cell lines and to evaluate the expression of key MMR proteins (MLH1, MSH2, MSH6, PMS2) to elucidate molecular differences between MSI and microsatellite stable (MSS) phenotypes. A panel of CRC cell lines with known MSI and MSS statuses was used. Telomere length was quantified using real-time quantitative PCR (qPCR) based on the T/S ratio method. MSI status was confirmed via PCR using mononucleotide repeat markers. Western blotting was performed to assess protein expressions of MLH1, MSH2, MSH6, and PMS2. β-actin served as a loading control. qPCR analysis revealed that MSI cell lines exhibited significantly longer telomeres compared to MSS lines (P < 0.05). Western blot results showed complete loss of MSH6 protein expression and reduction in expression levels of PMS2 and MSH2 in MSI cell lines, confirming MMR deficiency. In contrast, MSS cell lines maintained normal expression of all tested MMR proteins. These findings suggest a link between defective MMR function and altered telomere dynamics in MSI-CRC. MSI CRC cell lines exhibit telomere elongation and loss of key MMR proteins, highlighting distinct molecular features compared to MSS counterparts. These insights may inform future strategies for personalized CRC diagnostics and therapeutics, particularly in the context of telomere-targeted or immunomodulatory treatments.
This is an Open Access article, distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution and reproduction in any medium or format, provided the original work is properly cited.
Colorectal cancer (CRC) ranks as the third most common cancer globally and is the second leading cause of cancer-related mortality, with an estimated 1.9 million new cases and 935,000 deaths in 2020
[1]
Siegel RL, Miller KD, Fuchs HE, Jemal A. Cancer statistics, 2022. CA Cancer J Clin. 2022; 72: 7-33.
. Although CRC primarily affects individuals over the age of 50, an alarming rise in early-onset CRC has been noted in younger populations. The disease generally evolves from benign adenomatous or serrated polyps, progressing via a multi-step carcinogenic sequence driven by cumulative genetic and epigenetic alterations. The adenoma-carcinoma sequence, first conceptualized by Fearon and Vogelstein, continues to serve as a foundational framework for elucidating CRC pathogenesis
[2]
Fearon ER, Vogelstein B. A genetic model for colorectal tumorigenesis. Cell. 1990; 61(5): 759-767.
A hallmark of CRC is genomic instability, which manifests through three major and often overlapping mechanisms: chromosomal instability (CIN), microsatellite instability (MSI), and the CpG island methylator phenotype (CIMP)
[3]
Yamagishi H, Kuroda H, Imai Y, Hiraishi H. Molecular pathogenesis of sporadic colorectal cancers. Chin J Cancer. 2016; 35: 4.
Microsatellite instability (MSI) is defined by the accumulation of length variations in short tandem repeats due to defective DNA mismatch repair (MMR)
[4]
Boland CR, Goel A. Microsatellite instability in colorectal cancer. Gastroenterology. 2010; 138(6): 2073-2087. e3.
. First reported in CRC in the early 1990s, MSI is present in approximately 15-20% of sporadic CRCs and more than 95% of Lynch syndrome-associated tumors
[4]
Boland CR, Goel A. Microsatellite instability in colorectal cancer. Gastroenterology. 2010; 138(6): 2073-2087. e3.
Umar A, Boland CR, Terdiman JP, et al. Revised Bethesda Guidelines for hereditary nonpolyposis colorectal cancer (Lynch syndrome) and microsatellite instability. J Natl Cancer Inst. 2004; 96(4): 261-268.
. MSI is classified into three categories—MSI-high (MSI-H), MSI-low (MSI-L), and microsatellite stable (MSS)—based on the frequency and pattern of instability
[5]
Umar A, Boland CR, Terdiman JP, et al. Revised Bethesda Guidelines for hereditary nonpolyposis colorectal cancer (Lynch syndrome) and microsatellite instability. J Natl Cancer Inst. 2004; 96(4): 261-268.
. MSI-H tumors are associated with high tumor mutational burden (TMB), enhanced neoantigen presentation, better prognosis, and improved response to immune checkpoint inhibitors
[6]
Dey A, Mitra A, Pathak S, Prasad S, Zhang AS, Zhang H, et al. Recent advancements, limitations, and future perspectives of the use of personalized medicine in treatment of colon cancer. Technol Cancer Res Treat. 2023; 22: 15330338231178404.
The MMR pathway is vital for maintaining genomic fidelity by correcting base-base mismatches and insertion-deletion loops during DNA replication. Core MMR proteins include MLH1, PMS2, MSH2, and MSH6, which operate as heterodimers (e.g., MutSα, MutLα) to recognize and repair replication errors
[7]
Li GM. Mechanisms and functions of DNA mismatch repair. Cell Res. 2008; 18(1): 85-98.
Telomeres, composed of tandem TTAGGG repeats, protect chromosomal termini from degradation and fusion. Progressive telomere shortening, a natural consequence of cellular replication, can induce senescence or apoptosis
[10]
de Lange T. Shelterin: the protein complex that shapes and safeguards human telomeres. Genes Dev. 2018; 32(7-8): 527-540.
. In CRC, telomere attrition contributes to chromosomal instability and tumor evolution. MSI-H tumors have been shown to possess shorter telomeres than MSS counterparts, suggesting a correlation between MMR deficiency and telomere dynamics
[11]
Takagi S, Kinouchi Y, Hiwatashi N, et al. Relationship between microsatellite instability and telomere shortening in colorectal cancer. Jpn J Cancer Res. 2000; 1(4): 399-407.
. Moreover, loss of MMR proteins such as MLH1 and MSH2 promotes homologous recombination at telomeres, potentially enabling the alternative lengthening of telomeres (ALT) pathway
[12]
Bozzao C, Lastella P, Ponz de Leon M, Pedroni M, Di Gregorio C, D’Ovidio FD, et al. Analysis of telomere dynamics in peripheral blood cells from patients with Lynch syndrome. Cancer. 2011; 117: 4325-4335.
Telomerase, a ribonucleoprotein complex composed of the catalytic subunit hTERT and the RNA component hTERC, is reactivated in nearly 90% of human cancers, including CRC
[13]
Armanios M, Blackburn EH. The telomere syndromes. Nat Rev Genet. 2012; 13(10): 693-704.
. Intriguingly, MSI-H CRCs frequently exhibit diminished hTERT expression, indicating potential reliance on ALT for telomere maintenance
[15]
Omori Y, Nakayama F, Li D, Kanemitsu K, Semba S, Ito A, et al. Alternative lengthening of telomeres frequently occurs in mismatch repair system-deficient gastric carcinoma. Cancer Sci. 2009; 100: 413-418.
. Therefore, exploring the interactions between MMR deficiency and telomere regulation may offer novel insights into CRC biology and therapeutic targets.
2. Materials and Methods
2.1. Cell Culture
Human CRC cell lines CACO2 (MSS, RRID: CVCL_0025) and HCT15 (MSI, RRID: CVCL_0292) were procured and cultured under standard conditions at 37°C in a humidified 5% CO₂ atmosphere. CACO2 cells were maintained in Dulbecco’s Modified Eagle Medium (DMEM) supplemented with 20% fetal bovine serum (FBS), while HCT15 cells were grown in RPMI 1640 medium with 10% FBS. All protocols adhered to ATCC guidelines
[16]
Freshney RI. Culture of Animal Cells: A Manual of Basic Technique and Specialized Applications. 6th ed. Wiley-Liss; 2010.
Cells were harvested at ~90% confluence using trypsin-EDTA. Genomic DNA was extracted by phenol-chloroform method
[17]
Sambrook J, Russell DW. Molecular Cloning: A Laboratory Manual. 3rd ed. Cold Spring Harbor Laboratory Press; 2001.
[17]
, verified for integrity via 1% agarose gel electrophoresis, and quantified using a NanoDrop 2000 spectrophotometer (Thermo Fisher Scientific). Protein extraction was performed using RIPA buffer, followed by quantification and analysis according to standard protocols
[18]
Harlow E, Lane D. Using Antibodies: A Laboratory Manual. Cold Spring Harbor Laboratory Press; 2006.
[18]
.
2.3. Telomere Length Measurement
Telomere length was assessed using monochrome multiplex quantitative real-time PCR (MMqPCR) as described by Cawthon
[20]
Vasilishina A, Kropotov A, Spivak I, Bernadotte A. Relative human telomere length quantification by real-time PCR. Methods Mol Biol. 2019; 1973: 39-44.
Raji N, Majidi Zadeh T, Babheidarian P, Houshmand M. Detection of Microsatellite Instability by High-Resolution Melting Analysis in Colorectal Cancer. Iran Biomed J. 2022 Jan 1; 26(1): 70-6.
Hosen I, Rachakonda PS, Heidenreich B, de Verdier PJ, Ryk C, Steineck G, et al. Mutations in TERT promoter and FGFR3 and telomere length in bladder cancer. Int J Cancer. 2015; 137: 1621-1629.
Reactions were performed using the QuantStudio 6 Pro Real-Time PCR system. T/S ratios (telomere repeat copy number to single-copy gene number) were calculated from Ct values to estimate relative telomere length
[20]
Vasilishina A, Kropotov A, Spivak I, Bernadotte A. Relative human telomere length quantification by real-time PCR. Methods Mol Biol. 2019; 1973: 39-44.
MSI status was determined using high-resolution melting (HRM) analysis adapted from Raji et al
[21]
Raji N, Majidi Zadeh T, Babheidarian P, Houshmand M. Detection of Microsatellite Instability by High-Resolution Melting Analysis in Colorectal Cancer. Iran Biomed J. 2022 Jan 1; 26(1): 70-6.
. The pentaplex panel included five mononucleotide markers
[22]
Suraweera N, Duval A, Reperant M, et al. Evaluation of tumor microsatellite instability using five quasi-monomorphic mononucleotide repeats and pentaplex PCR. Oncogene. 2002; 21(37): 6041-6047.
. Lysates were electrophoresed on SDS-PAGE and transferred to membranes via semi-dry transfer. Membranes were blocked with 5% BSA or milk, then probed overnight at 4°C with primary antibodies against hMLH1, hMSH2, hMSH6, hPMS2, and β-actin
[25]
Jiricny J. The multifaceted mismatch-repair system. Nat Rev Mol Cell Biol. 2006; 7(5): 335-346.
. HRP-conjugated secondary antibodies were used, and bands were visualized using DAB or luminol substrate. Images were captured using ImageQuant software.
3. Results
3.1. DNA Quality Assessment and Quantification
For the MSI HCT15 cell line, the DNA showed high purity with A260/A280 ratios ranging between 1.80 and 1.90, and DNA yields were consistently above 100 ng/µL. Similarly, MSS CACO2 DNA samples demonstrated excellent integrity, with no smearing observed on gels and comparable purity metrics. These results confirmed that the genomic DNA was of suitable quality for downstream applications such as qPCR and HRM analyses.
3.2. Telomere Length Estimation
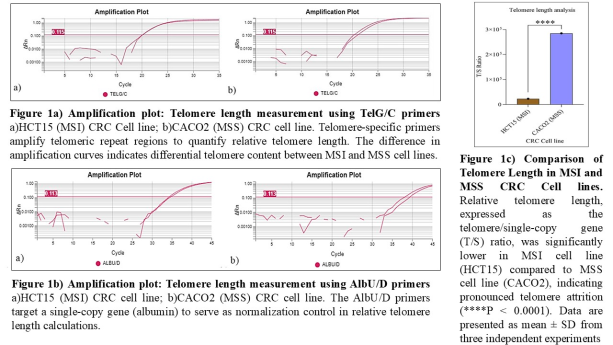
MMqPCR analysis revealed a significantly reduced telomere length (T/S ratio) in MSI-H HCT15 cells compared to MSS CACO2 cells (p < 0.05). The mean T/S ratio in HCT15 was 0.71 ± 0.08, whereas CACO2 cells exhibited a higher T/S ratio of 1.26 ± 0.11 (Figure 1). This indicates accelerated telomere shortening in MSI-H cells, aligning with the hypothesis that MMR deficiency may contribute to telomere erosion.
Table 1. Telomere Length Estimation in CRC Cell Lines via T/S Ratio Relative telomere length (T/S ratio) in HCT15 (MSI) and CACO2 (MSS) cell lines was determined using the ΔCt method (CtT - CtS) and calculated as 2ΔCt. Ct values were obtained via qPCR using TelG/C (telomeric) and AlbU/D (single-copy) primers. SD reflects variability across replicates.
Cell Line
CtT (TelG/C)
CtS (AlbU/D)
T/S Ratio (2ΔCt)
SD
HCT15 (MSI)
20.13
34.61
22,862.18
1.28
CACO2 (MSS)
20.60
38.72
284,641.89
1.63
3.3. Microsatellite Instability Analysis
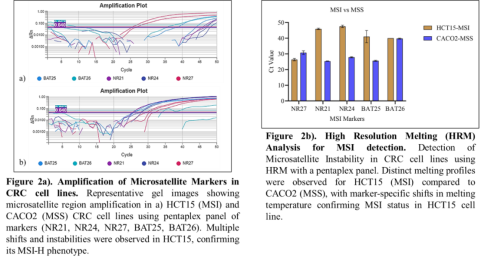
HRM profiles generated from the five-marker pentaplex assay (BAT-25, BAT-26, NR-21, NR-24, and NR-27) showed significant heteroduplex formation and distinct melting curve shifts in HCT15, confirming its MSI-H status. In contrast, CACO2 cells showed stable melting profiles across all five markers, consistent with an MSS phenotype (Figure 2). These findings validate the suitability of HRM as a rapid, dye-based, high-throughput method for MSI detection in CRC cell lines.
Table 2. MSI Marker Amplification in CRC Cell Lines by HRM-qPCR Mean Ct values and SDs for five MSI markers analyzed via HRM-qPCR in HCT15 (MSI) and CACO2 (MSS) cells. Ct shifts indicate differential amplification efficiency related to MSI status.
MSI Marker
HCT15 (MSI) - Mean Ct
SD
CACO2 (MSS) - Mean Ct
SD
NR27
26.37
0.77
30.80
1.18
NR21
45.97
0.46
25.42
0.08
NR24
47.58
0.75
27.87
0.35
BAT25
41.05
3.95
25.62
0.37
BAT26
40.00
0.00
39.76
0.35
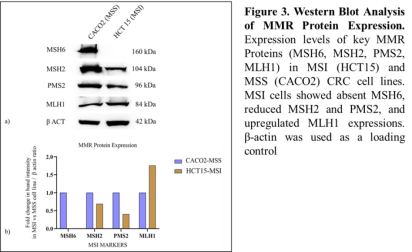
3.4. Western Blotting of MMR Proteins
Western blot analysis revealed a complete loss of MSH6 protein expression in the MSI-H HCT15 cell line, whereas MSH6 was robustly expressed in the MSS CACO2 cells. The expression levels of MSH2 and PMS2 were also significantly reduced in HCT15 cells, indicating downstream destabilization of the MMR heterodimers due to the absence of MSH6. Interestingly, MLH1 expression showed a 1.7-fold increase in the MSI cell line, suggesting a potential compensatory response. β-actin expression remained consistent across both cell lines, confirming equal protein loading (Figure 3).
Table 3. Protein Quantification in CRC Cell Lysates via BCA Assay Total protein concentrations (μg/mL) in CACO2 and HCT15 lysates were determined using the BCA assay. Absorbance at 562 nm was blank-corrected and values calculated using a standard curve. Measurements were performed in duplicate.
Colorectal cancer (CRC) pathogenesis involves diverse molecular subtypes, with microsatellite instability (MSI) and telomere dysfunction being pivotal contributors. This study demonstrates a significant correlation between MSI status and telomere shortening, coupled with downregulation of DNA mismatch repair (MMR) proteins, suggesting a mechanistic interplay between genomic instability and telomere biology in CRC. Our findings contribute to a growing body of evidence implicating the involvement of MSI and telomere dysfunction in CRC development.
MMR Deficiency and MSI in CRC: Our HRM-based MSI assay effectively differentiated between MSI-H (HCT15) and MSS (CACO2) phenotypes. This was in agreement with previous reports that highlight the critical role of MMR proteins in maintaining genomic stability
[26]
Martinez P, Siegl‐Cachedenier I, Flores JM, Blasco MA. MSH2 deficiency abolishes the anticancer and pro‐aging activity of short telomeres. Aging Cell. 2009; 8(1): 2-17.
Mendez‐Bermudez A, Royle NJ. Deficiency in DNA mismatch repair increases the rate of telomere shortening in normal human cells. Hum Mutat. 2011; 32(8): 939-946.
. The analysis revealed that MSH6 was not expressed in the MSI cell line, and there was downregulation of MSH2 and PMS2. Conversely, MLH1 expression was upregulated. These results suggest that the MSI cell line's heightened mutational burden and disrupted telomere maintenance may be due to the compromised functionality of specific MMR proteins
[28]
Vilar E, Gruber SB. Microsatellite instability in colorectal cancer—the stable evidence. Nat Rev Clin Oncol. 2010; 7(3): 153-162.
Kinouchi Y, Hiwatashi N, Chida M, Nagashima F, Takagi S, Maekawa H, et al. Telomere shortening in the colonic mucosa of patients with ulcerative colitis. J Gastroenterol. 1998; 33: 343–348.
. In particular, this study highlights that the absence of MSH6 protein expression is a significant contributor to microsatellite instability (MSI) in the CRC cell line. This deficiency not only induces MSI due to the inability to repair MMR mismatches but also appears to play a crucial role in the accelerated telomere shortening observed in MSI CRC cell lines. The simultaneous loss or reduced expression of MSH6, PMS2, and MSH2 likely compromises DNA repair efficiency, thereby exacerbating genomic instability. The elevated MLH1 expression in the MSI cell line may indicate a compensatory response to the loss of other MMR proteins, although its precise role in this context requires further investigation. The general classification and characterization of MSI across various types of cancer have been systematically outlined in previous studies, and de la Chapelle (2003) provided the foundational understanding of MSI’s role in CRC
[30]
de la Chapelle A. Microsatellite instability. N Engl J Med. 2003; 349(3): 209-210.
Telomere Shortening in MSI-H CRC: Our data on telomere length reveals a significant reduction in telomere size in HCT15 cells, in agreement with previous studies suggesting that MSI-H tumors tend to harbor shorter telomeres compared to their MSS counterparts
[31]
Denchi EL. Give me a break: How telomeres suppress the DNA damage response. DNA Repair (Amst). 2009;8(9):1118–1126.
Nersisyan L, Hopp L, Loeffler-Wirth H, Galle J, Loeffler M, Arakelyan A, et al. Telomere length maintenance and its transcriptional regulation in Lynch syndrome and sporadic colorectal carcinoma. Front Oncol. 2019; 9: 1172.
. This reduction in telomere length could be attributed to the loss of MMR function, which normally prevents aberrant homologous recombination at telomeres. The absence of functional MMR proteins likely leads to the destabilization of telomeres, promoting telomere attrition in MSI-H tumors. Moreover, the observed reduction in telomere length aligns with the hypothesis that MSI-H tumors may rely on alternative telomere lengthening (ALT) mechanisms in the absence of telomerase reactivation, a phenomenon well-documented in MMR-deficient cancers
[33]
Greider CW, Blackburn EH. Identification of a specific telomere terminal transferase activity in Tetrahymena extracts. Cell. 1985; 43(2 Pt 1): 405-413.
. These findings align with the systematic analysis of telomere lengths and somatic alterations in CRC, which reveal a close association between telomere attrition and tumor progression
[30]
de la Chapelle A. Microsatellite instability. N Engl J Med. 2003; 349(3): 209-210.
Therapeutic and Diagnostic Implications: The findings of this study support the growing paradigm that MSI status and telomere dynamics can be leveraged for CRC prognosis and therapy. MSI-H tumors, due to their high neo-antigen burden, are known to respond favorably to immune checkpoint blockade therapies
[34]
Barthel FP, Wei W, Tang M, et al. Systematic analysis of telomere length and somatic alterations in 31 cancer types. Nat Genet. 2017; 49(3): 349-357.
. Understanding the interplay between telomere dysfunction and MSI may open new avenues for targeted therapeutic strategies. For example, telomere-based diagnostics could be utilized to stratify patients for telomerase-targeted or ALT-inhibitory therapies, both of which may hold promises for improving clinical outcomes in MSI-H CRC patients.
Limitations and Future Directions: While this study provides robust in vitro data, it is limited to only two cell lines, which may not fully capture the complexity of CRC heterogeneity. Future work should aim to include primary tumor samples from MSI-H and MSS CRC patients to validate these observations and assess their clinical relevance. Additionally, telomere length assays, telomerase activity assays, and ALT pathway analyses could provide further insights into the telomere maintenance strategies employed by different CRC subtypes. Expanding the study to encompass a broader range of CRC models will help to define the full extent of telomere dysfunction and its therapeutic implications in MSI-associated cancers.
Abbreviations
CRC
Colorectal Cancer
MSI
Microsatellite Instability
MSI-H
Microsatellite Instability - High
MMR
Mismatch Repair
MSS
Microsatellite Stable
CIN
Chromosomal Instability
CIMP
CpG island Methylator Phenotype
ALT
Alternative Lengthening of Telomeres
MMqPCR
Monochrome Multiplex Quantitative Real-Time PCR
T/S Ratio
Telomere Repeat Copy Number to Single-Copy Gene Number
HRM
High-Resolution Melting
Acknowledgments
The authors gratefully acknowledge the Manipal Academy of Higher Education (MAHE) and the Manipal School of Life Sciences for providing essential infrastructure and institutional support throughout the course of this research. Special thanks are extended to the TIFAC-CORE in Pharmacogenomics, MAHE, for facilitating access to advanced research facilities.
Author Contributions
Rajasekhar Moka: Conceptualization, Writing-Review and Editing, Supervision.
Umar A, Boland CR, Terdiman JP, et al. Revised Bethesda Guidelines for hereditary nonpolyposis colorectal cancer (Lynch syndrome) and microsatellite instability. J Natl Cancer Inst. 2004; 96(4): 261-268.
Dey A, Mitra A, Pathak S, Prasad S, Zhang AS, Zhang H, et al. Recent advancements, limitations, and future perspectives of the use of personalized medicine in treatment of colon cancer. Technol Cancer Res Treat. 2023; 22: 15330338231178404.
Takagi S, Kinouchi Y, Hiwatashi N, et al. Relationship between microsatellite instability and telomere shortening in colorectal cancer. Jpn J Cancer Res. 2000; 1(4): 399-407.
Bozzao C, Lastella P, Ponz de Leon M, Pedroni M, Di Gregorio C, D’Ovidio FD, et al. Analysis of telomere dynamics in peripheral blood cells from patients with Lynch syndrome. Cancer. 2011; 117: 4325-4335.
Omori Y, Nakayama F, Li D, Kanemitsu K, Semba S, Ito A, et al. Alternative lengthening of telomeres frequently occurs in mismatch repair system-deficient gastric carcinoma. Cancer Sci. 2009; 100: 413-418.
Sambrook J, Russell DW. Molecular Cloning: A Laboratory Manual. 3rd ed. Cold Spring Harbor Laboratory Press; 2001.
[18]
Harlow E, Lane D. Using Antibodies: A Laboratory Manual. Cold Spring Harbor Laboratory Press; 2006.
[19]
Hosen I, Rachakonda PS, Heidenreich B, de Verdier PJ, Ryk C, Steineck G, et al. Mutations in TERT promoter and FGFR3 and telomere length in bladder cancer. Int J Cancer. 2015; 137: 1621-1629.
Vasilishina A, Kropotov A, Spivak I, Bernadotte A. Relative human telomere length quantification by real-time PCR. Methods Mol Biol. 2019; 1973: 39-44.
Raji N, Majidi Zadeh T, Babheidarian P, Houshmand M. Detection of Microsatellite Instability by High-Resolution Melting Analysis in Colorectal Cancer. Iran Biomed J. 2022 Jan 1; 26(1): 70-6.
Suraweera N, Duval A, Reperant M, et al. Evaluation of tumor microsatellite instability using five quasi-monomorphic mononucleotide repeats and pentaplex PCR. Oncogene. 2002; 21(37): 6041-6047.
Martinez P, Siegl‐Cachedenier I, Flores JM, Blasco MA. MSH2 deficiency abolishes the anticancer and pro‐aging activity of short telomeres. Aging Cell. 2009; 8(1): 2-17.
Mendez‐Bermudez A, Royle NJ. Deficiency in DNA mismatch repair increases the rate of telomere shortening in normal human cells. Hum Mutat. 2011; 32(8): 939-946.
Kinouchi Y, Hiwatashi N, Chida M, Nagashima F, Takagi S, Maekawa H, et al. Telomere shortening in the colonic mucosa of patients with ulcerative colitis. J Gastroenterol. 1998; 33: 343–348.
Nersisyan L, Hopp L, Loeffler-Wirth H, Galle J, Loeffler M, Arakelyan A, et al. Telomere length maintenance and its transcriptional regulation in Lynch syndrome and sporadic colorectal carcinoma. Front Oncol. 2019; 9: 1172.
Moka, R., Upadhya, S. (2025). Telomere Elongation and Mismatch Repair Deficiency Distinguish MSI Colorectal Cancer Cell Lines. Cancer Research Journal, 13(2), 23-29. https://doi.org/10.11648/j.crj.20251302.11
Moka R, Upadhya S. Telomere Elongation and Mismatch Repair Deficiency Distinguish MSI Colorectal Cancer Cell Lines. Cancer Res J. 2025;13(2):23-29. doi: 10.11648/j.crj.20251302.11
@article{10.11648/j.crj.20251302.11,
author = {Rajasekhar Moka and Shreya Upadhya},
title = {Telomere Elongation and Mismatch Repair Deficiency Distinguish MSI Colorectal Cancer Cell Lines
},
journal = {Cancer Research Journal},
volume = {13},
number = {2},
pages = {23-29},
doi = {10.11648/j.crj.20251302.11},
url = {https://doi.org/10.11648/j.crj.20251302.11},
eprint = {https://article.sciencepublishinggroup.com/pdf/10.11648.j.crj.20251302.11},
abstract = {Microsatellite instability (MSI) is a hallmark of mismatch repair (MMR) deficiency and characterizes a distinct subset of colorectal cancers (CRC). In parallel, telomere length dynamics have emerged as important contributors to genomic stability and tumorigenesis. However, the relationship between MSI status, MMR protein expression, and telomere maintenance remains poorly defined. This study aimed to investigate the association between MSI status and telomere length in CRC cell lines and to evaluate the expression of key MMR proteins (MLH1, MSH2, MSH6, PMS2) to elucidate molecular differences between MSI and microsatellite stable (MSS) phenotypes. A panel of CRC cell lines with known MSI and MSS statuses was used. Telomere length was quantified using real-time quantitative PCR (qPCR) based on the T/S ratio method. MSI status was confirmed via PCR using mononucleotide repeat markers. Western blotting was performed to assess protein expressions of MLH1, MSH2, MSH6, and PMS2. β-actin served as a loading control. qPCR analysis revealed that MSI cell lines exhibited significantly longer telomeres compared to MSS lines (P < 0.05). Western blot results showed complete loss of MSH6 protein expression and reduction in expression levels of PMS2 and MSH2 in MSI cell lines, confirming MMR deficiency. In contrast, MSS cell lines maintained normal expression of all tested MMR proteins. These findings suggest a link between defective MMR function and altered telomere dynamics in MSI-CRC. MSI CRC cell lines exhibit telomere elongation and loss of key MMR proteins, highlighting distinct molecular features compared to MSS counterparts. These insights may inform future strategies for personalized CRC diagnostics and therapeutics, particularly in the context of telomere-targeted or immunomodulatory treatments.
},
year = {2025}
}
TY - JOUR
T1 - Telomere Elongation and Mismatch Repair Deficiency Distinguish MSI Colorectal Cancer Cell Lines
AU - Rajasekhar Moka
AU - Shreya Upadhya
Y1 - 2025/05/29
PY - 2025
N1 - https://doi.org/10.11648/j.crj.20251302.11
DO - 10.11648/j.crj.20251302.11
T2 - Cancer Research Journal
JF - Cancer Research Journal
JO - Cancer Research Journal
SP - 23
EP - 29
PB - Science Publishing Group
SN - 2330-8214
UR - https://doi.org/10.11648/j.crj.20251302.11
AB - Microsatellite instability (MSI) is a hallmark of mismatch repair (MMR) deficiency and characterizes a distinct subset of colorectal cancers (CRC). In parallel, telomere length dynamics have emerged as important contributors to genomic stability and tumorigenesis. However, the relationship between MSI status, MMR protein expression, and telomere maintenance remains poorly defined. This study aimed to investigate the association between MSI status and telomere length in CRC cell lines and to evaluate the expression of key MMR proteins (MLH1, MSH2, MSH6, PMS2) to elucidate molecular differences between MSI and microsatellite stable (MSS) phenotypes. A panel of CRC cell lines with known MSI and MSS statuses was used. Telomere length was quantified using real-time quantitative PCR (qPCR) based on the T/S ratio method. MSI status was confirmed via PCR using mononucleotide repeat markers. Western blotting was performed to assess protein expressions of MLH1, MSH2, MSH6, and PMS2. β-actin served as a loading control. qPCR analysis revealed that MSI cell lines exhibited significantly longer telomeres compared to MSS lines (P < 0.05). Western blot results showed complete loss of MSH6 protein expression and reduction in expression levels of PMS2 and MSH2 in MSI cell lines, confirming MMR deficiency. In contrast, MSS cell lines maintained normal expression of all tested MMR proteins. These findings suggest a link between defective MMR function and altered telomere dynamics in MSI-CRC. MSI CRC cell lines exhibit telomere elongation and loss of key MMR proteins, highlighting distinct molecular features compared to MSS counterparts. These insights may inform future strategies for personalized CRC diagnostics and therapeutics, particularly in the context of telomere-targeted or immunomodulatory treatments.
VL - 13
IS - 2
ER -
Moka, R., Upadhya, S. (2025). Telomere Elongation and Mismatch Repair Deficiency Distinguish MSI Colorectal Cancer Cell Lines. Cancer Research Journal, 13(2), 23-29. https://doi.org/10.11648/j.crj.20251302.11
Moka R, Upadhya S. Telomere Elongation and Mismatch Repair Deficiency Distinguish MSI Colorectal Cancer Cell Lines. Cancer Res J. 2025;13(2):23-29. doi: 10.11648/j.crj.20251302.11
@article{10.11648/j.crj.20251302.11,
author = {Rajasekhar Moka and Shreya Upadhya},
title = {Telomere Elongation and Mismatch Repair Deficiency Distinguish MSI Colorectal Cancer Cell Lines
},
journal = {Cancer Research Journal},
volume = {13},
number = {2},
pages = {23-29},
doi = {10.11648/j.crj.20251302.11},
url = {https://doi.org/10.11648/j.crj.20251302.11},
eprint = {https://article.sciencepublishinggroup.com/pdf/10.11648.j.crj.20251302.11},
abstract = {Microsatellite instability (MSI) is a hallmark of mismatch repair (MMR) deficiency and characterizes a distinct subset of colorectal cancers (CRC). In parallel, telomere length dynamics have emerged as important contributors to genomic stability and tumorigenesis. However, the relationship between MSI status, MMR protein expression, and telomere maintenance remains poorly defined. This study aimed to investigate the association between MSI status and telomere length in CRC cell lines and to evaluate the expression of key MMR proteins (MLH1, MSH2, MSH6, PMS2) to elucidate molecular differences between MSI and microsatellite stable (MSS) phenotypes. A panel of CRC cell lines with known MSI and MSS statuses was used. Telomere length was quantified using real-time quantitative PCR (qPCR) based on the T/S ratio method. MSI status was confirmed via PCR using mononucleotide repeat markers. Western blotting was performed to assess protein expressions of MLH1, MSH2, MSH6, and PMS2. β-actin served as a loading control. qPCR analysis revealed that MSI cell lines exhibited significantly longer telomeres compared to MSS lines (P < 0.05). Western blot results showed complete loss of MSH6 protein expression and reduction in expression levels of PMS2 and MSH2 in MSI cell lines, confirming MMR deficiency. In contrast, MSS cell lines maintained normal expression of all tested MMR proteins. These findings suggest a link between defective MMR function and altered telomere dynamics in MSI-CRC. MSI CRC cell lines exhibit telomere elongation and loss of key MMR proteins, highlighting distinct molecular features compared to MSS counterparts. These insights may inform future strategies for personalized CRC diagnostics and therapeutics, particularly in the context of telomere-targeted or immunomodulatory treatments.
},
year = {2025}
}
TY - JOUR
T1 - Telomere Elongation and Mismatch Repair Deficiency Distinguish MSI Colorectal Cancer Cell Lines
AU - Rajasekhar Moka
AU - Shreya Upadhya
Y1 - 2025/05/29
PY - 2025
N1 - https://doi.org/10.11648/j.crj.20251302.11
DO - 10.11648/j.crj.20251302.11
T2 - Cancer Research Journal
JF - Cancer Research Journal
JO - Cancer Research Journal
SP - 23
EP - 29
PB - Science Publishing Group
SN - 2330-8214
UR - https://doi.org/10.11648/j.crj.20251302.11
AB - Microsatellite instability (MSI) is a hallmark of mismatch repair (MMR) deficiency and characterizes a distinct subset of colorectal cancers (CRC). In parallel, telomere length dynamics have emerged as important contributors to genomic stability and tumorigenesis. However, the relationship between MSI status, MMR protein expression, and telomere maintenance remains poorly defined. This study aimed to investigate the association between MSI status and telomere length in CRC cell lines and to evaluate the expression of key MMR proteins (MLH1, MSH2, MSH6, PMS2) to elucidate molecular differences between MSI and microsatellite stable (MSS) phenotypes. A panel of CRC cell lines with known MSI and MSS statuses was used. Telomere length was quantified using real-time quantitative PCR (qPCR) based on the T/S ratio method. MSI status was confirmed via PCR using mononucleotide repeat markers. Western blotting was performed to assess protein expressions of MLH1, MSH2, MSH6, and PMS2. β-actin served as a loading control. qPCR analysis revealed that MSI cell lines exhibited significantly longer telomeres compared to MSS lines (P < 0.05). Western blot results showed complete loss of MSH6 protein expression and reduction in expression levels of PMS2 and MSH2 in MSI cell lines, confirming MMR deficiency. In contrast, MSS cell lines maintained normal expression of all tested MMR proteins. These findings suggest a link between defective MMR function and altered telomere dynamics in MSI-CRC. MSI CRC cell lines exhibit telomere elongation and loss of key MMR proteins, highlighting distinct molecular features compared to MSS counterparts. These insights may inform future strategies for personalized CRC diagnostics and therapeutics, particularly in the context of telomere-targeted or immunomodulatory treatments.
VL - 13
IS - 2
ER -