Background: Chronic exposure to aluminium chloride (AlCl3) damages the hippocampus and impairs cognition through oxidative stress, reactive astrogliosis, and neuronal apoptosis. Phytochemicals with antioxidant and anti-inflammatory activity are increasingly considered as candidate neuroprotectants, yet evidence supporting combined plant-derived interventions that target these convergent pathways remains scarce. Purpose: This study examined whether Kolaviron, a biflavonoid complex from Garcinia kola seeds, and an ethanolic extract of Bryophyllum pinnatum (CRA), administered individually or in combination, attenuate AlCl3-induced hippocampal injury and cognitive impairment in Wistar rats, and whether their protective actions converge on the Nrf2 signalling pathway. Methods: Seventy adult male Wistar rats were randomly assigned to seven groups of ten: vehicle control, AlCl3 (100 mg/kg), Kolaviron alone (200 mg/kg), CRA alone (600 mg/kg), AlCl3 + Kolaviron, AlCl3 + CRA, and AlCl3 + Kolaviron + CRA. All agents were administered by oral gavage daily for fourteen days. Spatial learning and memory were assessed using the Morris water maze. Hippocampal homogenates were assayed for superoxide dismutase (SOD), catalase (CAT), and malondialdehyde (MDA). Haematoxylin and eosin staining was used to evaluate cytoarchitecture, and immunohistochemistry quantified glial fibrillary acidic protein (GFAP), B-cell lymphoma 2 (Bcl-2), and nuclear factor erythroid 2-related factor 2 (Nrf2) expression in the CA3 subfield. Data were analysed by one-way analysis of variance (ANOVA) followed by Tukey’s post hoc test. Results: AlCl3 exposure prolonged escape latency, lowered SOD and CAT activities, raised MDA, produced neuronal loss with pyknotic and vacuolated cells, increased GFAP immunoreactivity, and reduced both Bcl-2 and Nrf2 expression in CA3. Co-treatment with Kolaviron or CRA reversed each of these alterations (p < 0.05 versus AlCl3 alone), and the combined regimen produced the most consistent restoration across behavioural, biochemical, histological, and immunohistochemical endpoints, frequently returning values close to control levels. Conclusion: Kolaviron and B. pinnatum protect the rat hippocampus against AlCl3-induced damage by restoring antioxidant defences, attenuating astrogliosis, preserving Bcl-2 expression, and activating Nrf2 signalling, with the combination conferring broader protection than either agent alone. These findings support further investigation of these phytochemicals as candidates against environmental neurotoxicant-induced neurodegeneration.
| Published in | Clinical Neurology and Neuroscience (Volume 10, Issue 2) |
| DOI | 10.11648/j.cnn.20261002.12 |
| Page(s) | 60-70 |
| Creative Commons |
This is an Open Access article, distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution and reproduction in any medium or format, provided the original work is properly cited. |
| Copyright |
Copyright © The Author(s), 2026. Published by Science Publishing Group |
Aluminum, Oxidative Stress, Nrf2 Pathway, Hippocampus, Kolaviron
AD | Alzheimer’s Disease |
AlCl3 | Aluminium Chloride |
ANOVA | Analysis of Variance |
ARE | Antioxidant Response Element |
ARRIVE | Animal Research Reporting of In Vivo Experiments |
Bcl-2 | B-cell lymphoma 2 |
CA3 | Cornu Ammonis 3 (Hippocampal Subfield) |
CAT | Catalase |
CRA | Crassulaceae (Bryophyllum Pinnatum Ethanolic Extract) |
DAB | 3,3′-Diaminobenzidine |
GFAP | Glial Fibrillary Acidic Protein |
H&E | Haematoxylin and Eosin |
HRP | Horseradish Peroxidase |
Keap1 | Kelch-like ECH-associated Protein 1 |
KV | Kolaviron |
MDA | Malondialdehyde |
MWM | Morris Water Maze |
NF-κB | Nuclear Factor Kappa B |
NIH | National Institutes of Health |
Nrf2 | Nuclear Factor Erythroid 2-related Factor 2 |
PD | Parkinson’s Disease |
ROS | Reactive Oxygen Species |
SEM | Standard Error of the Mean |
SOD | Superoxide Dismutase |
TBARS | Thiobarbituric Acid Reactive Substances |
| [1] | Nabi M, Tabassum N. Role of environmental toxicants in neurodegenerative disorders. Frontiers in Toxicology. 2022; 4: 837579. |
| [2] | Adelodun ST, Ishola OA, Abijo AZ, Olatunji SY, Owolabi JO, Olanrewaju JA, Adekomi DA. Aluminium chloride-induced hippocampal damage: CA3 hippocampal subfield involvement and the neuroprotective role of Buchholzia coriacea ethanolic seed extract. Phytomedicine Plus. 2021; 1(3): 100104. |
| [3] | Liaquat L, Sadir S, Batool Z, Tabassum S, Shahzad S, Afzal A, Haider S. Acute aluminium chloride toxicity revisited: Study on DNA damage and histopathological, biochemical and neurochemical alterations in rat brain. Life Sciences. 2019; 217: 202–211. |
| [4] | Bhargava VP, Netam AK, Singh R, Sharma P. Cassia tora mitigates aluminium chloride-induced alterations in pro-inflammatory cytokines, neurotransmitters, and beta-amyloid and tau protein markers in Wistar rats. Toxicology International. 2023; 30(1): 63–81. |
| [5] | Dash UC, Bhol NK, Swain SK, Samal RR, Nayak PK, Raina V, Panda SK, Kerry RG, Duttaroy AK, Jena AB. Oxidative stress and inflammation in the pathogenesis of neurological disorders: Mechanisms and implications. Acta Pharmaceutica Sinica B. 2024; 15(1): 15–34. |
| [6] | Jin C, Li Y, Chai J, Zhang Y, Li Y. Aluminium induces neuroinflammation via the P2X7 receptor activating the NLRP3 inflammasome pathway. Ecotoxicology and Environmental Safety. 2023; 249: 114373. |
| [7] | Nazari-Serenjeh M, Baluchnejadmojarad T, Hatami-Morassa M, Fahanik-Babaei J, Mehrabi S, Roghani M. Kolaviron neuroprotective effect against okadaic acid-provoked cognitive impairment. Heliyon. 2024; 10(3): e025564. |
| [8] | Crapser JD, Spangenberg EE, Barahona RA, Arreola MA, Hohsfield LA, Green KN. Microglia facilitate loss of perineuronal nets in the Alzheimer’s disease brain. eBioMedicine. 2020; 58: 102919. |
| [9] | Adamu A, Li S, Gao F, Xue G. The role of neuroinflammation in neurodegenerative diseases: Current understanding and future therapeutic targets. Frontiers in Aging Neuroscience. 2024; 16: 1347987. |
| [10] | Abdel-Aal RA, Hussein OA, Elsaady RG, Abdelzaher LA. Naproxen as a potential candidate for promoting rivastigmine anti-Alzheimer activity against aluminium chloride-prompted Alzheimer’s-like disease in rats: Neurogenesis and apoptosis modulation as a possible underlying mechanism. European Journal of Pharmacology. 2022; 915: 174695. |
| [11] | Cuadrado A, Rojo AI, Wells G, Hayes JD, Cousin SP, Rumsey WL, Attucks OC, Franklin G, Levonen AL, Kensler TW, Dinkova-Kostova AT. Therapeutic targeting of the NRF2-KEAP1 partnership in chronic diseases. Nature Reviews Drug Discovery. 2019; 18(4): 295–317. |
| [12] | Andrade EA de, Machinski I, Ventura ACT, Barr SA, Pereira AV, Beltrame FL, Strangman WK, Williamson RT. A review of the popular uses, anatomical, chemical, and biological aspects of Kalanchoe (Crassulaceae): A genus of plants known as “Miracle Leaf.” Molecules. 2023; 28(14): 5574. |
| [13] | Awogbindin IO, Onasanwo SA, Ezekiel OO, Akindoyeni I, Mustapha Y, Farombi OE. Neuroprotection of kolaviron by regulation of nuclear factor erythroid 2-related factor 2 in an MPTP mice model of Parkinson’s disease. American Journal of Biopharmacy and Pharmaceutical Sciences. 2021; 1(1): 5. |
| [14] | Ogidigo JO, Anosike CA, Joshua PE, Ibeji CU, Nwanguma BC, Nwodo OFC. Neuroprotective effect of Bryophyllum pinnatum flavonoids against aluminium chloride-induced neurotoxicity in rats. Toxicology Mechanisms and Methods. 2022; 32(4): 243–258. |
| [15] | Farombi EO, Abarikwu SO, Adedara IA, Oyeyemi MO. Curcumin and kolaviron ameliorate di-n-butylphthalate-induced testicular damage in rats. Basic & Clinical Pharmacology & Toxicology. 2007; 100(1): 43–48. |
| [16] | Campos HM, Costa MD, Moreira LK da S, Neri HF da S, Silva CRB da, Pruccoli L, Santos FCA dos, Costa ÉA, Tarozzi A, Ghedini PC. Protective effects of chrysin against the neurotoxicity induced by aluminium: In vitro and in vivo studies. Toxicology. 2021; 465: 153033. |
| [17] | Oyenihi OR, Brooks NL, Oguntibeju OO. Effects of kolaviron on hepatic oxidative stress in streptozotocin-induced diabetes. BMC Complementary and Alternative Medicine. 2015; 15: 236. |
| [18] | Nunez J. Morris water maze experiment. Journal of Visualized Experiments. 2008; (19): e897. |
| [19] | Ebokaiwe AP, Ramesh P, Mathur PP, Farombi EO. Transient effect of single dose exposure of Nigerian Bonny-light crude oil on testicular steroidogenesis in Wistar rats is accompanied by oxidative stress. Drug and Chemical Toxicology. 2015; 38(4): 428–435. |
| [20] | Akinrinade ID, Memudu AE, Ogundele OM, Ajetunmobi OI. Interplay of glia activation and oxidative stress formation in fluoride and aluminium exposure. Pathophysiology. 2015; 22(1): 39–48. |
| [21] | Erukainure OL, Ijomone OM, Sanni O, Aschner M, Islam M. Type 2 diabetes induced oxidative brain injury involves altered cerebellar neuronal integrity and elemental distribution, and exacerbated Nrf2 expression: Therapeutic potential of raffia palm (Raphia hookeri) wine. Metabolic Brain Disease. 2019; 34(5): 1385–1399. |
| [22] | Ijomone OM, Olatunji SY, Owolabi JO, Naicker T, Aschner M. Nickel-induced neurodegeneration in the hippocampus, striatum and cortex; an ultrastructural insight, and the role of caspase-3 and α-synuclein. Journal of Trace Elements in Medicine and Biology. 2018; 50: 16–23. |
| [23] | Ijomone OM, Nwoha PU. Nicotine inhibits hippocampal and striatal acetylcholinesterase activities, and demonstrates dual action on adult neuronal proliferation and maturation. Pathophysiology. 2015; 22(4): 231–239. |
| [24] | Misra HP, Fridovich I. The role of superoxide anion in the auto-oxidation of epinephrine and a simple assay for superoxide dismutase. Journal of Biological Chemistry. 1972; 247(10): 3170–3175. |
| [25] | Claiborne A. Catalase activity. In: Greenwald AR, editor. Handbook of methods for oxygen radical research. CRC Press; 1995. p. 237–242. |
| [26] | Aboelwafa HR, El-Kott AF, Abd-Ella EM, Yousef HN. The Possible Neuroprotective Effect of Silymarin against Aluminum Chloride-Prompted Alzheimer's-Like Disease in Rats. Brain Sci. 2020; 10(9): 628. |
| [27] | Onasanwo SA, Velagapudi R, El-Bakoush A, Olajide OA. Inhibition of neuroinflammation in BV2 microglia by the biflavonoid kolaviron is dependent on the Nrf2/ARE antioxidant protective mechanism. Mol Cell Biochem. 2016; 414(1-2): 23-36. |
| [28] | Bittencourt LO, Damasceno-Silva RD, Aragão WAB, Eiró-Quirino L, Oliveira ACA, Fernandes RM, Freire MAM, Cartágenes SC, Dionizio A, Buzalaf MAR, Cassoli JS, Cirovic A, Cirovic A, Maia CDSF, Lima RR. Global Proteomic Profile of Aluminum-Induced Hippocampal Impairments in Rats: Are Low Doses of Aluminum Really Safe? Int J Mol Sci. 2022; 23(20): 12523. |
| [29] | Guzman SJ, Schlögl A, Frotscher M, Jonas P. Synaptic mechanisms of pattern completion in the hippocampal CA3 network. Science. 2016; 353(6304): 1117-1123. |
| [30] | Zhang Z, Li X, Ma L, Wang S, Zhang J, Zhou Y, Guo X, Niu Q. LNC000152 Mediates Aluminum-Induced Proliferation of Reactive Astrocytes. ACS Omega. 2024; 9(10): 11958-11968. |
| [31] | Arndt H, Bachurski M, Yuanxiang P, Franke K, Wessjohann LA, Kreutz MR, Grochowska KM. A Screen of Plant-Based Natural Products Revealed That Quercetin Prevents Pyroglutamylated Amyloid-β (Aβ3(pE)-42) Uptake in Astrocytes As Well As Resulting Astrogliosis and Synaptic Dysfunction. Mol Neurobiol. 2025; 62(3): 3730-3745. |
| [32] | Brandes MS, Gray NE. NRF2 as a therapeutic target in neurodegenerative diseases. ASN Neuro. 2020; 12: 1759091419899782. |
| [33] | Dewanjee S, Bhattacharya H, Bhattacharyya C, Chakraborty P, Fleishman J, Alexiou A, Papadakis M, Jha SK. Nrf2/Keap1/ARE regulation by plant secondary metabolites: A new horizon in brain tumor management. Cell Communication and Signaling. 2024; 22: 497. |
| [34] | Niture SK, Jaiswal AK. Nrf2 protein up-regulates antiapoptotic protein Bcl-2 and prevents cellular apoptosis. Journal of Biological Chemistry. 2012; 287(13): 9873–9886. |
| [35] | Costa LG, Garrick JM, Roquè PJ, Pellacani C. Mechanisms of neuroprotection by quercetin: counteracting oxidative stress and more. Oxidative Medicine and Cellular Longevity. 2016; 2016: 2986796. |
APA Style
Oria, R. S., Mesole, B. S., Otu, U. G., Akpang, K. E., Usin, S. G., et al. (2026). Kolaviron and Bryophyllum pinnatum Attenuate AlCl3-Induced Memory Impairment by Modulating Oxidative Stress, Astrogliosis, and Bcl-2/Nrf2 Signaling. Clinical Neurology and Neuroscience, 10(2), 60-70. https://doi.org/10.11648/j.cnn.20261002.12
ACS Style
Oria, R. S.; Mesole, B. S.; Otu, U. G.; Akpang, K. E.; Usin, S. G., et al. Kolaviron and Bryophyllum pinnatum Attenuate AlCl3-Induced Memory Impairment by Modulating Oxidative Stress, Astrogliosis, and Bcl-2/Nrf2 Signaling. Clin. Neurol. Neurosci. 2026, 10(2), 60-70. doi: 10.11648/j.cnn.20261002.12
AMA Style
Oria RS, Mesole BS, Otu UG, Akpang KE, Usin SG, et al. Kolaviron and Bryophyllum pinnatum Attenuate AlCl3-Induced Memory Impairment by Modulating Oxidative Stress, Astrogliosis, and Bcl-2/Nrf2 Signaling. Clin Neurol Neurosci. 2026;10(2):60-70. doi: 10.11648/j.cnn.20261002.12
@article{10.11648/j.cnn.20261002.12,
author = {Rademene Sunday Oria and Bolaji Samuel Mesole and Ujong Gabriel Otu and Kingsley Ekpe Akpang and Saviour God’swealth Usin and Cynthia Nyen Tangban and Sesugh Ugese and Comfort Andokie Ugi Ugbong and Chidera Michael Nwodo and Oluchi Priscilla Uwazurike and Chinecherem Marvelous Obidinanwa and Margaret Ikanobi Michael},
title = {Kolaviron and Bryophyllum pinnatum Attenuate
AlCl3-Induced Memory Impairment by Modulating Oxidative Stress, Astrogliosis, and Bcl-2/Nrf2 Signaling},
journal = {Clinical Neurology and Neuroscience},
volume = {10},
number = {2},
pages = {60-70},
doi = {10.11648/j.cnn.20261002.12},
url = {https://doi.org/10.11648/j.cnn.20261002.12},
eprint = {https://article.sciencepublishinggroup.com/pdf/10.11648.j.cnn.20261002.12},
abstract = {Background: Chronic exposure to aluminium chloride (AlCl3) damages the hippocampus and impairs cognition through oxidative stress, reactive astrogliosis, and neuronal apoptosis. Phytochemicals with antioxidant and anti-inflammatory activity are increasingly considered as candidate neuroprotectants, yet evidence supporting combined plant-derived interventions that target these convergent pathways remains scarce. Purpose: This study examined whether Kolaviron, a biflavonoid complex from Garcinia kola seeds, and an ethanolic extract of Bryophyllum pinnatum (CRA), administered individually or in combination, attenuate AlCl3-induced hippocampal injury and cognitive impairment in Wistar rats, and whether their protective actions converge on the Nrf2 signalling pathway. Methods: Seventy adult male Wistar rats were randomly assigned to seven groups of ten: vehicle control, AlCl3 (100 mg/kg), Kolaviron alone (200 mg/kg), CRA alone (600 mg/kg), AlCl3 + Kolaviron, AlCl3 + CRA, and AlCl3 + Kolaviron + CRA. All agents were administered by oral gavage daily for fourteen days. Spatial learning and memory were assessed using the Morris water maze. Hippocampal homogenates were assayed for superoxide dismutase (SOD), catalase (CAT), and malondialdehyde (MDA). Haematoxylin and eosin staining was used to evaluate cytoarchitecture, and immunohistochemistry quantified glial fibrillary acidic protein (GFAP), B-cell lymphoma 2 (Bcl-2), and nuclear factor erythroid 2-related factor 2 (Nrf2) expression in the CA3 subfield. Data were analysed by one-way analysis of variance (ANOVA) followed by Tukey’s post hoc test. Results: AlCl3 exposure prolonged escape latency, lowered SOD and CAT activities, raised MDA, produced neuronal loss with pyknotic and vacuolated cells, increased GFAP immunoreactivity, and reduced both Bcl-2 and Nrf2 expression in CA3. Co-treatment with Kolaviron or CRA reversed each of these alterations (p 3 alone), and the combined regimen produced the most consistent restoration across behavioural, biochemical, histological, and immunohistochemical endpoints, frequently returning values close to control levels. Conclusion: Kolaviron and B. pinnatum protect the rat hippocampus against AlCl3-induced damage by restoring antioxidant defences, attenuating astrogliosis, preserving Bcl-2 expression, and activating Nrf2 signalling, with the combination conferring broader protection than either agent alone. These findings support further investigation of these phytochemicals as candidates against environmental neurotoxicant-induced neurodegeneration.},
year = {2026}
}
TY - JOUR T1 - Kolaviron and Bryophyllum pinnatum Attenuate AlCl3-Induced Memory Impairment by Modulating Oxidative Stress, Astrogliosis, and Bcl-2/Nrf2 Signaling AU - Rademene Sunday Oria AU - Bolaji Samuel Mesole AU - Ujong Gabriel Otu AU - Kingsley Ekpe Akpang AU - Saviour God’swealth Usin AU - Cynthia Nyen Tangban AU - Sesugh Ugese AU - Comfort Andokie Ugi Ugbong AU - Chidera Michael Nwodo AU - Oluchi Priscilla Uwazurike AU - Chinecherem Marvelous Obidinanwa AU - Margaret Ikanobi Michael Y1 - 2026/06/18 PY - 2026 N1 - https://doi.org/10.11648/j.cnn.20261002.12 DO - 10.11648/j.cnn.20261002.12 T2 - Clinical Neurology and Neuroscience JF - Clinical Neurology and Neuroscience JO - Clinical Neurology and Neuroscience SP - 60 EP - 70 PB - Science Publishing Group SN - 2578-8930 UR - https://doi.org/10.11648/j.cnn.20261002.12 AB - Background: Chronic exposure to aluminium chloride (AlCl3) damages the hippocampus and impairs cognition through oxidative stress, reactive astrogliosis, and neuronal apoptosis. Phytochemicals with antioxidant and anti-inflammatory activity are increasingly considered as candidate neuroprotectants, yet evidence supporting combined plant-derived interventions that target these convergent pathways remains scarce. Purpose: This study examined whether Kolaviron, a biflavonoid complex from Garcinia kola seeds, and an ethanolic extract of Bryophyllum pinnatum (CRA), administered individually or in combination, attenuate AlCl3-induced hippocampal injury and cognitive impairment in Wistar rats, and whether their protective actions converge on the Nrf2 signalling pathway. Methods: Seventy adult male Wistar rats were randomly assigned to seven groups of ten: vehicle control, AlCl3 (100 mg/kg), Kolaviron alone (200 mg/kg), CRA alone (600 mg/kg), AlCl3 + Kolaviron, AlCl3 + CRA, and AlCl3 + Kolaviron + CRA. All agents were administered by oral gavage daily for fourteen days. Spatial learning and memory were assessed using the Morris water maze. Hippocampal homogenates were assayed for superoxide dismutase (SOD), catalase (CAT), and malondialdehyde (MDA). Haematoxylin and eosin staining was used to evaluate cytoarchitecture, and immunohistochemistry quantified glial fibrillary acidic protein (GFAP), B-cell lymphoma 2 (Bcl-2), and nuclear factor erythroid 2-related factor 2 (Nrf2) expression in the CA3 subfield. Data were analysed by one-way analysis of variance (ANOVA) followed by Tukey’s post hoc test. Results: AlCl3 exposure prolonged escape latency, lowered SOD and CAT activities, raised MDA, produced neuronal loss with pyknotic and vacuolated cells, increased GFAP immunoreactivity, and reduced both Bcl-2 and Nrf2 expression in CA3. Co-treatment with Kolaviron or CRA reversed each of these alterations (p 3 alone), and the combined regimen produced the most consistent restoration across behavioural, biochemical, histological, and immunohistochemical endpoints, frequently returning values close to control levels. Conclusion: Kolaviron and B. pinnatum protect the rat hippocampus against AlCl3-induced damage by restoring antioxidant defences, attenuating astrogliosis, preserving Bcl-2 expression, and activating Nrf2 signalling, with the combination conferring broader protection than either agent alone. These findings support further investigation of these phytochemicals as candidates against environmental neurotoxicant-induced neurodegeneration. VL - 10 IS - 2 ER -
Department of Human Anatomy, University of Cross River State, Yala, Nigeria
Human Anatomy Unit, Eden University, Lusaka, Zambia
Department of Human Physiology, University of Cross River State, Yala, Nigeria
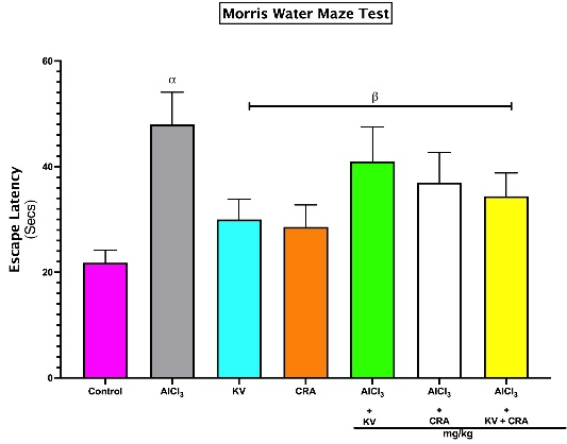
Figure 1. Escape Latency of rats exposed to Aluminium Chloride (AlCl3) and treated with Kolaviron and/or Crassulaceae extract. Bars represent mean ± SEM; n=5. α indicates significant difference from control (p<0.05); β indicates significant difference from AlCl3 only group (p<0.05). One-way ANOVA followed by Tukey post-hoc test. (AlCl3 - Aluminium Chloride, KV - Kolaviron, CRA - Crassulaceae).
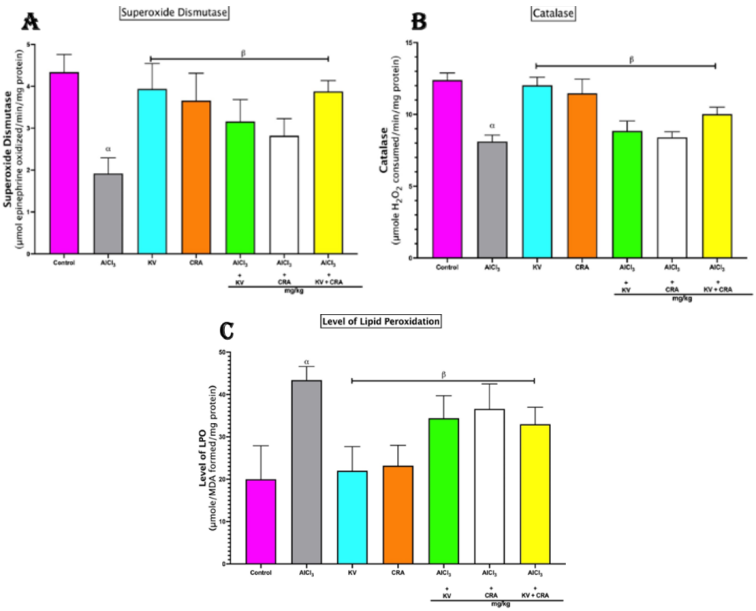
Figure 2. Influence of Kolaviron and/or Crassulaceae extract on antioxidant status and lipid peroxidation in the brain tissue of rats exposed to Aluminium Chloride (AlCl3). Bars represent mean ± SEM; n=5. α indicates significant difference from control (p<0.05); β indicates significant difference from AlCl3 only group (p<0.05). One-way ANOVA followed by Tukey post-hoc test. (AlCl3 - Aluminium Chloride, KV - Kolaviron, CRA - Crassulaceae).
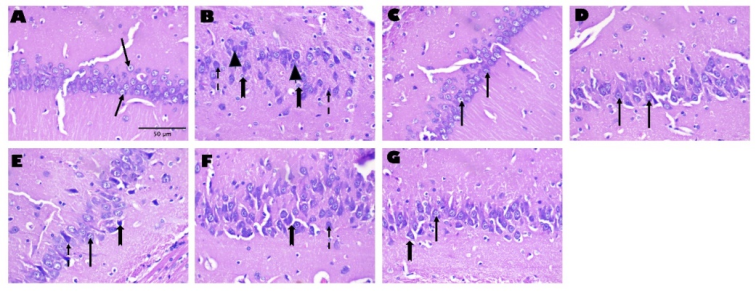
Figure 3. Hematoxylin and eosin (H&E) staining of the hippocampus (CA3) in rat brain tissue sections following exposure to Aluminium Chloride (AlCl3) and treatment with Kolaviron and/or Crassulaceae extract. Magnification: 400×; Scale bars: 50μm. Arrows: intact neurons; Dashed arrow: neurons with prominent eosinophilic cytoplasm; Notched arrows: neuronal swelling and/or vacuolation; Ạrrow head: Pyknotic nuclei.
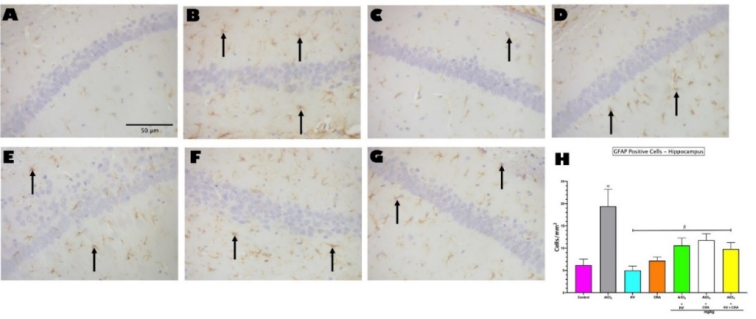
Figure 4. Photomicrograph showing immunohistochemical staining of GFAP-positive Cells in the hippocampus (A-G). Magnification: 400 x; Scale bars: 50μm. Black arrows indicate GFAP-positive Astrocytes. H shows Image J analysis of GFAP-positive cells in the Hippocampus (CA3). Bars represent mean ± SEM; n=5. α indicates significant difference from control (p<0.05); β indicates significant difference from AlCl3 only group (p<0.05). One-way ANOVA followed by Tukey post-hoc test. (AlCl3 - Aluminium Chloride, KV - Kolaviron, CRA - Crassulaceae).
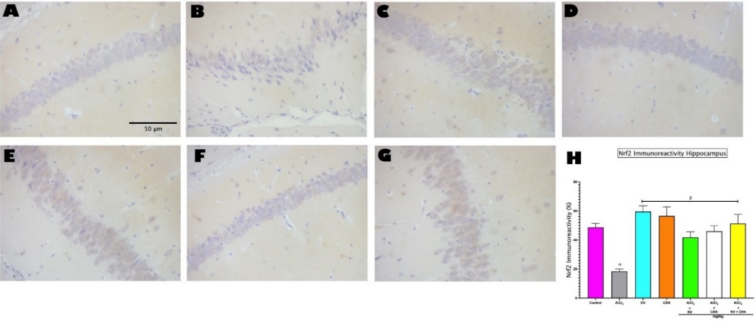
Figure 5. Photomicrograph showing Immunoreactivity of Nrf2 in the hippocampus (A-G). Magnification: 400 ×; Scale bars: 50μm. H shows Image J analysis of Nrf2 Immunoreactivity in the Hippocampus (CA3). Bars represent mean ± SEM; n=5. α indicates significant difference from control (p<0.05); β indicates significant difference from AlCl3 only group (p<0.05). One-way ANOVA followed by Tukey post-hoc test. (AlCl3 - Aluminium Chloride, KV - Kolaviron, CRA - Crassulaceae).
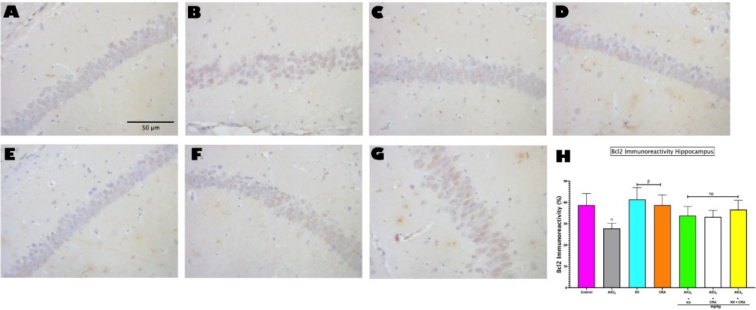
Figure 6. Photomicrograph showing Immunoreactivity of Bcl2 in the hippocampus (A-G). Magnification: 400 ×; Scale bars: 50μm. H shows Image J analysis of Bcl2 Immunoreactivity in the Hippocampus (CA3). Bars represent mean ± SEM; n=5. α indicates significant difference from control (p<0.05); β indicates significant difference from AlCl3 only group (p<0.05). One-way ANOVA followed by Tukey post-hoc test. (AlCl3 - Aluminium Chloride, KV - Kolaviron, CRA - Crassulaceae).
Information

